Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
Aminosyrenedbryting og ureaproduksjon
R-CH-COO- NH3+ Winnie Eskild, IMBV 2004

2
Katabolisme av aminosyrer
Aminosyrer som brytes ned kommer fra 3 kilder: Overskudd av inntak fra proteiner. Proteinene spaltes først i aminosyrer som deretter nedbrytes. Kroppen kan ikke lagre ekstra proteiner Daglig utskifting av proteiner Muskel som brytes ned i forbindelse med sult eller sykdom, diabetes

3
Katabolisme av aminosyrer
De fleste aminosyrer brukes på nytt Som karbohydrater og fett kan også størsteparten av en aminosyre brytes ned i det vevet som vil bruke den til energiproduksjon Aminosyrenes karbonskjelett kan enten oksideres til CO2 og H2O eller det kan brukes som utgangspunkt for syntese av glukose via glukoneogenese reaksjonsveien

4
Katabolisme av aminosyrer
Aminosyrenes aminogruppe brytes ned i spesielle reaksjoner i leveren Kroppen har derfor en transportoppgave når det gjelder å få aminogrupper fra alle perifert degraderte aminosyrer til leveren

5
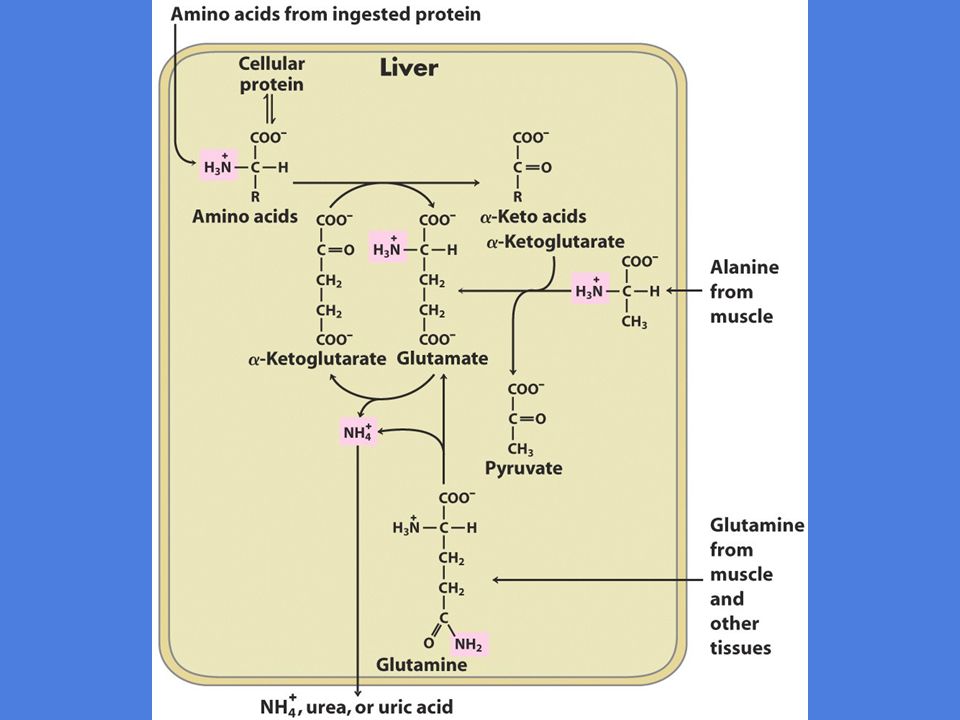
Oversikt over aminosyre katabolismen

6
Aminogruppens metabolske skjebne
Nitrogen er vanskelig å håndtere i metabolismen fordi det er for inert for biokjemiske prosesser Organismen tar derfor vare på og gjenutnytter sine aminogrupper Tre aminosyrer spiller en spesiell rolle i aminosyremetabolismen: Glutamat, glutamin og alanin I levercellenes cytosol overføres de fleste aminogrupper til a-ketoglutarat som omdannes til glutamat

7
Aminogruppens metabolske skjebne
Glutamat transporteres inn i mitokondriet hvor aminogruppen metaboliseres I de fleste andre vev overføres aminogruppen til glutamat som omdannes til glutamin Glutamin transporteres til leveren hvor aminogruppen metaboliseres

8
Aminogruppens metabolske skjebne
I muskel overføres aminogruppen til pyruvat som omdannes til alanin Alanin transporteres til leveren hvor aminogruppen metaboliseres

10
Nitrogen utskilles på tre måter
NH4+ utskilles av fisk, amfibier. Vannet de lever i løser det opp Urea utskilles av vertebrater, haier. Krever en del vann, skånsomt for vev Urinsyre utskilles av fugler, krypdyr. Krystalliserer, veier lite og krever ikke vann Ammonoteliske organismer Ureoteliske organismer Urikoteliske organismer

11
Samling av NH2-grupper i glutamat Transamineringsreaksjonen
Overføring av en aminogruppe fra et molekyle til et annet kalles en transaminering Reaksjonen katalyseres av transaminaser eller aminotransferaser Hver aminosyre har sin spesifikke aminotransferase som har navn etter den aminosyre som gir fra seg en aminogruppe i reaksjonen Eksempel: alanin aminotransferase overfører -NH2 fra alanin til a-ketoglutarat Aminotransferaser bruker pyridoksalfosfat som kofaktor

12
Pyridoksalfosfat Pyridoksalfosfat er kovalent bundet til en lysinsidekjede i enzymet, prostetisk gruppe Pyridoksalfosfat bærer amino-gruppen under reaksjonen

13
Pyridoksalfosfat Pyridoksalfosfat er en prostetisk gruppe
PLP er bundet med en kovalent dobbelbinding til en lysinsidekjede i enzymet, Schiff base Pyridoksalfosfat bærer amino-gruppen under reaksjonen Dobbelbindingen til enzymet erstattes av en enkelbinding til aminogruppen

14
Fri NH4+ er toksisk Noe fri NH4+ dannes bl. a. i hjernen
Enzymet glutamin syntetase katalyserer innkorporering av NH4+ i glutamin Glutamin sendes via blodsirkulasjonen til leveren. Glutamin er neutralt, ikke toksisk og passerer lett cellemembranen

15
Glutamin transporterer aminogrupper fra periferien til leveren
Glutamin transporteres inn i mitokondriet i lever eller nyre Enzymet glutaminase frigjør amid NH2-gruppen NH4+ går til ureasyklus Glutamat går inn i glutamat- poolen i mitokondriet

16
Oksidativ deaminering
Glutamat bærer aminogrupper fra mange aminosyrer i leveren Etter overføring til mitokondriet gjennomgår glutamat oksidativ deaminering Enzymet er glutamat dehydrogenase med NAD+/NADP+ som kofaktor a-ketoglutarat går til sitronsyresyklus eller glukoneogenese Allosterisk enzym, GTP hemmer, ADP stimulerer aktiviteten

17
Alanin som -NH2-transportør
Glukose-alanin syklus Arbeidende muskel har anaerob glykolyse => pyruvat kan ikke gi mere energi Pyruvat får overført en -NH2-gruppe og blir til alanin Føres med blodet til leveren hvor NH2-gruppen overføres til -ketoglutarat av enzymet alanin aminotransferase og pyruvat omdannes til glukose Glutamat overføres til mitokondriet hvor aminogruppen frigjøres av enzymet glutamat dehydrogenase Glukose sendes tilbake til muskelen

18
Samling av NH4+ Aminogrupper fra perifere vev og fra reaksjoner i cytosol frigjøres for å bli ført inn i ureasyklus

19
Samling av NH4+ Aminogrupper fra perifere vev og fra reaksjoner i cytosol frigjøres for å bli ført inn i ureasyklus

20
Ureasyklus Ureasyklus omdanner aminogrupper til urea
Prosessen involverer enzymer i mitokondriet såvel som i cytosol Omdannelsen av aminogrupper til urea starter i mitokondriet og sluttføres i cytosol Ureasyklus inngår i tett samspill med sitronsyresyklus. Dette samspillet kalles ofte for Krebs bisyklus (Sitronsyre syklus = Krebs syklus)
")
21
Syntese av karbamoylfosfat
Enzymet er karbamoylfosfat syntetase I, et allosterisk enzym, N-acetylglutamat stimulerer Reaksjonen har flere trinn og krever investering av 2 ATP CO2 + H2O <=> HCO3- + H+ Bikarbonat reagerer først med ATP og fører til dannelse av en karbonsyre-fosforsyre anhydrid

22
Syntese av karbamoylfosfat
Deretter spaltes anhydridbindingen og en ammoniumion koples på C-atomet. Vi får dannet karbamat

23
Syntese av karbamoylfosfat
Karbamat fosforyleres ved bruk av ATP og vi får dannet karbamoylfosfat som har en anhydrid-binding Karbamoylfosfat kan betraktes som aktivert NH4+

24
Første trinn i ureasyklus foregår i mitokondriet
Karbamoylfosfat overfører sin karbamoylgruppe til ornitin og det dannes citrullin. Pi frigis Reaksjon drives av hydrolysen av anhydridbindingen mellom karbamoylgruppen og fosfatgruppen Enzymet er ornitin transkarbamoylase Ornitin er en aminosyre, finnes bare som metabolismeintermediat Citrullin forlater mitokondriet

25
Cytosoliske trinn i ureasyklus
Citrullin til argininosuccinat Citrullin aktiveres ved påkopling av AMP på karbamoyl C-atomet Enzymet er argininosuccinat syntetase For å drive reksjonen fremover hydrolyseres pyrofosfat til 2 Pi 2 Pi

26
Cytosoliske trinn i ureasyklus
Aminogruppe nr. 2 skal nå hentes inn Enzymet er argininosuccinat syntetase Dette gjøres ved å bytte ut AMP med aspartat, som bindes via sin amino-gruppe til ureidogruppen i citrullin (det som var karbamoyl C-atomet) Det er nå dannet argininosuccinat
 Det er nå dannet argininosuccinat.")
27
Cytosoliske trinn i ureasyklus
Her spaltes argininosuccinat til arginin og fumarat Enzymet er argininosuccinat lyase

28
Cytosoliske trinn i ureasyklus
Fumarat går tilbake til mitokondriet Via sitronsyresyklus gjendannes det til oksaloacetat Oksaloacetat kan få overført en ny aminogruppe fra glutamat og derved bli omdannet til aspartat Aspartat forlater mitokondriet og reagerer med citrullin

29
Cytosoliske trinn i ureasyklus
Enzymet arginase spalter arginin i ornitin og urea Ornitin går tilbake inn i mitokondriet, klar til å hente en ny karbamoylgruppe Urea passerer over i blodet og utskilles av nyrene til urinen De cytosoliske enzymene i ureasyklus sitter tett assosiert på cytosolsiden av indre mitokondriemembran => substratkanalisering og dermed høy effektivitet

30
Ureasyklus

31
Krebs bisyklus Fumarase og malat dehydro-genase finnes som både cytosoliske og mitokondrielle isoenzymer

32
Regulering av ureasyklus
Proteiner brytes ned og aminosyrer forbrennes til energi etter inntak av overskudd av protein eller ved langvarig sult Den økte frisetting av aminogrupper øker hastigheten i ureasyklus Det fins korttids- og langtidsregulering for ureasyklus Langtidsregulering: her reguleres syntesen av ureasyklusenzymene i leveren Syntesehastigheten for disse enzymene er høy ved sult eller proteinrik kost, mens den lav hos planteetere Korttidsregulering: her reguleres aktiviteten til allerede syntetiserte enzymer Aktiviteten til karbamoylfosfat syntetase reguleres allosterisk

33
Reguleringav karbamoylfosfat syntetase I
Karbamoylfosfat syntetase I er allosterisk stimulert av N-acetyl-glutamat Ved høye nivåer av aminogrupper dannes N-acetylglutamat ved reaksjon mellom glutamat og acetyl-CoA Reaksjonen katalyseres av N-acetyl-glutamat syntase Dette enzymet er det første i reaksjons-veien som syntetiserer arginin fra glutamat Resten av enzymene i denne reaksjonsveien finnes ikke i leveren, bare i andre vev med aminosyre-syntese Reaksjonen tjener derfor bare reguleringsformål

34
Nettoreaksjon for ureasyntese
2 NH4+ + HCO ATP + H2O => urea + 2 ADP + 4 Pi + AMP + 5 H+ Det koster altså 4 ATP Fumarat kan kompensere dette noe. Omdannelse til oksaloacetat går via et oksidasjonstrinn og produserer en NADH. Når denne sender elektronene sine gjennom elektrontransportkjeden får vi dannet 2,5 ATP Nettokostnad for dannelse av urea er derfor nærmere 1,5 ATP

35
NH4+ er giftig Det er viktig å bli kvitt NH4+ på en kontrollert måte
NH4+ er en svak syre, mens NH3 er en relativt sterk base Alkanisering anses å være en del av problemet, dette er spesielt farlig for hjernen NH3 menes også å forstyrre hjernens ATP produksjon og syntesen av nevrotransmittere Assimilering av NH4+ inn i biomolekyler katalyseres bl.a. av glutamin syntetase som omdanner glutamat til glutamin. Dette enzymet har lav Km for NH4+ og finnes i alle organismer

Liknende presentasjoner