Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
Muskelkontraksjonens biokjemi 2015 Stadium 1B Hans E. Krokan og Magnus Steigedal, IKM

2
KILDER Hardin J, Bertoni G og Kleinsmith LJ. Becker’s World of the cell, eight ed. 2012 Mathews, Van Holde og Ahern. Biochemistry, third ed. 2000 Vander, Sherman og Luciano. Human Physiology, seventh ed. 1998 Germann WJ, Stanfield CL. Principles of Human Physiology. Benjamin Cummings 2001. Boron WF, Boulpaep EL. Medical Physiology. Saunders 2003. Lodish H, Berk A, Zipursky SL, Matsudaira P, Baltimore D, Darnell J. Molecular Cell Biology. WH Freeman and Company 2000. Martini FH. Fundamentals of Anatomy and Physiology. Benjamin Cummins. Meisenberg, G. and Simmons, W.H. Principles of Medical Biochemistry ELSEVIER Saunders 3 rd edition 2012

3
Læringsmål 5 Biokjemi, cellebiologi 5.1.2 gjøre rede for de biokjemiske mekanismene for muskelstimulering og -kontraksjon

4
MUSKELCELLER ER INTRIKATE “MASKINER” Har komponenter som tilsvarer dem vi finner hos neuroner (neuro-transmittor, reseptorer, spennings-sensitive ionekanaler osv.) og kan generere elektriske signaler som respons på nevrotransmitter Kan generere kraft og bevegelse på grunn av en rekke kontraktile proteiner som kan omforme kjemisk energi (fra hydrolyse av ATP) til mekanisk energi
 og kan generere elektriske signaler som respons på nevrotransmitter Kan generere kraft og bevegelse på grunn av en rekke kontraktile proteiner som kan omforme kjemisk energi (fra hydrolyse av ATP) til mekanisk energi")
5
KJENNETEGNES VED PRESISJON kan aktivere kontraksjonssystemet i løpet av millisekunder - og “ slå det av” på tilsvarende kort tid kan variere mengden kraft som mobiliseres - og tilpasse den til bevegelsen som skal utføres

6
TRE HOVEDTYPER MUSKULATUR SKJELETTMUSKULATUR HJERTEMUSKULATUR GLATT MUSKULATUR

7
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001

8
SKJELETTMUSKULATUR – FUNKSJONELLE ENHETER hierarkisk oppbygging

9
STRUKTUR MUSKELLEGEMET: den “kjøttaktige” del av muskelen FASCICLER: samling av individuelle muskelceller, bindevev, blodkar og nerver MUSKELCELLE/ FIBER: fusjonert celle med flere kjerner, strekker seg i muskelens fulle lengde SARCOLEMMA: muskelfiberens plasmamembran MYOFIBRILLER: stavformede elementer i muskelfibrenes cytoplasma, inneholder kontraksjonsmaskineriet

10
STRUKTUR forts. ACTIN & MYOSIN: proteiner som er bestanddeler i myofibrillene SARCOPLASMATISK RETICULUM: et sekkelignende membranøst nettverk som omgir hver av myofibrillene. Knyttes sammen ved hjelp av transverse tubuli. TRANSVERSE TUBULI: forbinder muskelfibrenes indre med sarcolemma (ytre)
.")
11
SARCOPLASMATISKE RETICULUM OG T-TUBULI overfører signaler fra sarcolemma til myofibrillene - gjør muskelcellen i stand til å respondere på et nevralt signal det sarcoplasmatiske reticulum lagrer Ca 2+ ioner og frigjør disse til cytosol når muskelcellen stimuleres til kontraksjon frigjøringen av Ca 2+ ioner skjer som resultat av elektriske signaler som går fra sarcolemma til T-tubuli

12
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001

15
Figure 16-76 Molecular Biology of the Cell (© Garland Science 2008) Sarcomer - den minste kontraktile enhet
 Sarcomer - den minste kontraktile enhet")
16
Myofibrillene består av repeterende enheter: SARCOMERER. Hver sarcomer begynner og slutter med en Z-linje (forbinder de tynne filamentene). De tykke filamentene er forbundet gjennom M-linjen.
. De tykke filamentene er forbundet gjennom M-linjen..")
17
Figure 16-74 Molecular Biology of the Cell (© Garland Science 2008) Elektronmikroskopisk (TME) bilde av muskelcelle
 Elektronmikroskopisk (TME) bilde av muskelcelle")
18
Figure 16-75 Molecular Biology of the Cell (© Garland Science 2008) TME bilde av muskelcelle – heksagonal struktur - to nivå av heksagonal (sekskantet) struktur
 TME bilde av muskelcelle – heksagonal struktur - to nivå av heksagonal (sekskantet) struktur")
19
Figure 16-76 Molecular Biology of the Cell (© Garland Science 2008) Sarcomer - den minste kontraktile enhet
 Sarcomer - den minste kontraktile enhet")
20
De tynne filamentene består av ACTIN. De tykke filamentene består av MYOSIN. Disse har tversgående utløpere –”kryssbindinger” (CROSSBRIDGES) som forbinder tykke og tynne filamenter.
 som forbinder tykke og tynne filamenter..")
21
Figure 16-54a Molecular Biology of the Cell (© Garland Science 2008) Myosin - bunter av disse bygger tykke filamenter
 Myosin - bunter av disse bygger tykke filamenter")
22
Figure 16-56b Molecular Biology of the Cell (© Garland Science 2008) Myosinhodet bindes til actin in vitro
 Myosinhodet bindes til actin in vitro")
23
Figure 16-57 Molecular Biology of the Cell (© Garland Science 2008) Myosin gener- en stor familie av proteinkodende gener De fleste klassene har flere, til dels mange ulike gener Type I: I alle celler – involvert i vesikkeltransport, monomer Type II: I skjelettmuskulatur (mange undertyper, bl.a. i hjerte) Type III-XIV: Dårlig kjent, eller ukjent funksjon Raske og langsomme fibre: dels pga. ulike typer myosin, dels avhengig av trening; fenotypene er i noen grad «interkonvertible».
 Type III-XIV: Dårlig kjent, eller ukjent funksjon Raske og langsomme fibre: dels pga. ulike typer myosin, dels avhengig av trening; fenotypene er i noen grad «interkonvertible»..")
24
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001 Myosinmolekyler – dimerer med spiralsstruktur Myosinmolekyler – sammenbindes hale-til-hale Myosinmolekyler – aggregerer slik at hoderekker danner 60 o vinkel med hverandre Mange myosinhoder bindes til hvert actin-filament Myosinhodet har ATPase domene som binder ATP og et actin-bindende domene som binder til actin

25
TYNNE FILAMENTER HAR I TILLEGG til actin to spesialiserte regulatoriske proteiner: TROPOMYOSIN: et langt, fibrøst molekyl som spenner over en rekke actinmolekyler slik at det blokkerer de myosinbindende setene i muskulatur i hvile TROPONIN er et kompleks av tre proteiner - ett binder til actin - ett binder til tropomyosin - ett binder til sete der Ca 2+ kan bindes (reversibelt)
")
26
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001 Actin subenheter er globulære molekyler (G actin) som interakterer og bygger et dobbelttrådet filament G-actin F-actin F-actin, tropomyosin og troponin-kompleks
 som interakterer og bygger et dobbelttrådet filament G-actin F-actin F-actin, tropomyosin og troponin-kompleks.")
27
TYKKE FILAMENTER består av myosin-molekyler (hundrevis). Hver enkelt er en dimer. HODET er “hodet” i myosinmolekylet. I denne delen genereres muskelens mekaniske kraft. - ett actinbindende sete - ett ATPase sete som har enzymatisk aktivitet og hydrolyserer ATP HODET danner “crossbridges” TITIN elastisk protein som kan strekkes til mer enn 3X sin naturlige lengde

28
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001 Myosinmolekyler – dimerer med spiralsstruktur Myosinmolekyler – sammenbindes hale-til-hale Myosinmolekyler – aggregerer slik at hoderekker danner 60 o vinkel med hverandre Mange myosinhoder bindes til hvert actin-filament Myosinhodet har ATPase domene som binder ATP og et actin-bindende domene som binder til actin

29
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001 Muskel kontraksjonen – krever ATP NB! Rigor mortis = dødsstivhet (situasjonen når all ATP er forbrukt)
.")
30
HVORDAN SKRUES MUSKELKONTRAKSJONER AV OG PÅ? Når en muskelcelle mottar signal fra et motorneuron, depolariserer cellen - og fyrer av et aksjonspotensial som i neste omgang stimulerer til kontraksjon. Sekvensen av hendelser kalles: EKSITASJONS - KONTRAKSJONSKOBLING

32
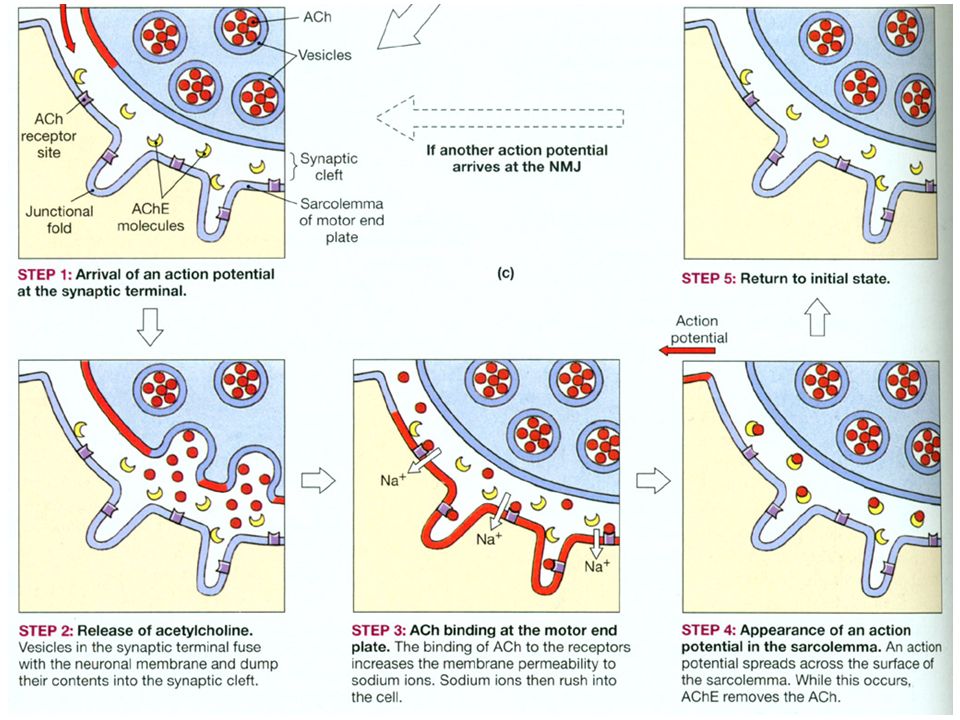
2. aksjonspotensialet vandrer langs sarcolemma og ned til T-tubuli. Bidrar til å frigjøre Ca 2+ fra sarcoplasmatisk reticulum. EKSITASJONS - KONTRAKSJONSKOBLING 1.motorneuronet overfører et aksjonspotensial og bidrar til utskillelse av neurotransmittoren acetylcholin. Acetylcholin binder seg til spesifikke reseptorer bidrar til endring i ionepermeabilitet endrer membranpotensialet depolarisering

33
Figure 16-77a Molecular Biology of the Cell (© Garland Science 2008)
")
34
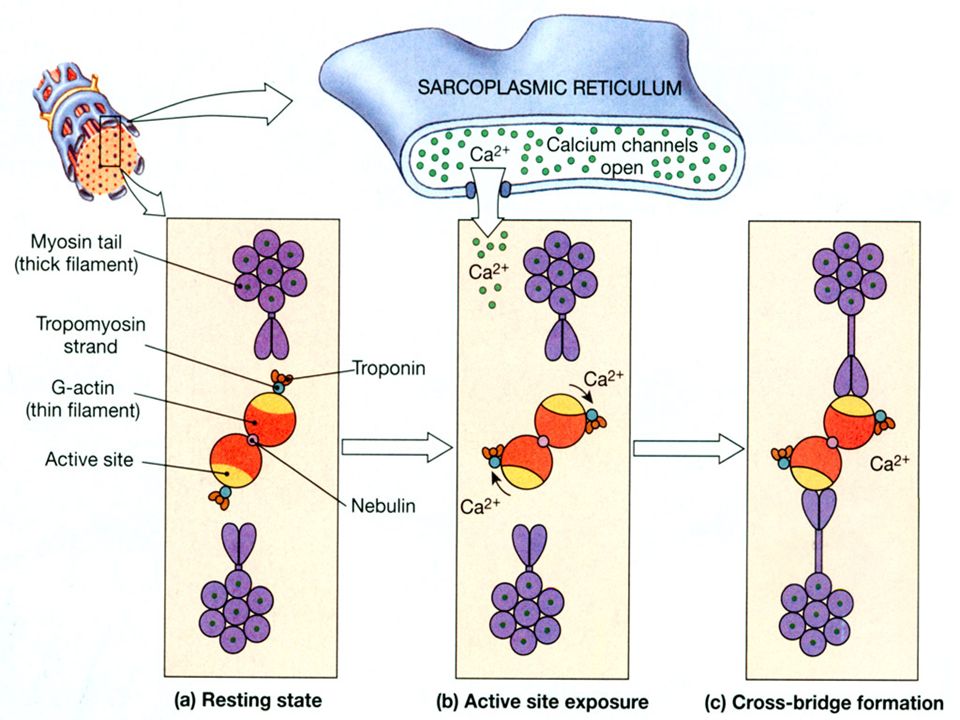
CA 2+, TROPONIN OG TROPOMYOSIN I EKSITASJONS-KONTRAKSJONS KOBLINGEN: når en muskel er avslappet, er konsentrasjonen av Ca 2+ i cytosol lav liten binding mellom Ca 2+ og troponin. Tropomyosin er posisjonert slik at actins myosinbindende seter er blokkert. når et aksjonspotensial i T-tubuli trigger frigjøring av Ca 2+, binder Ca 2+ seg til spesifikke bindingssteder i det sarcoplasmatiske reticulum disse åpnes mer Ca 2+ frigjøres

35
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001

37
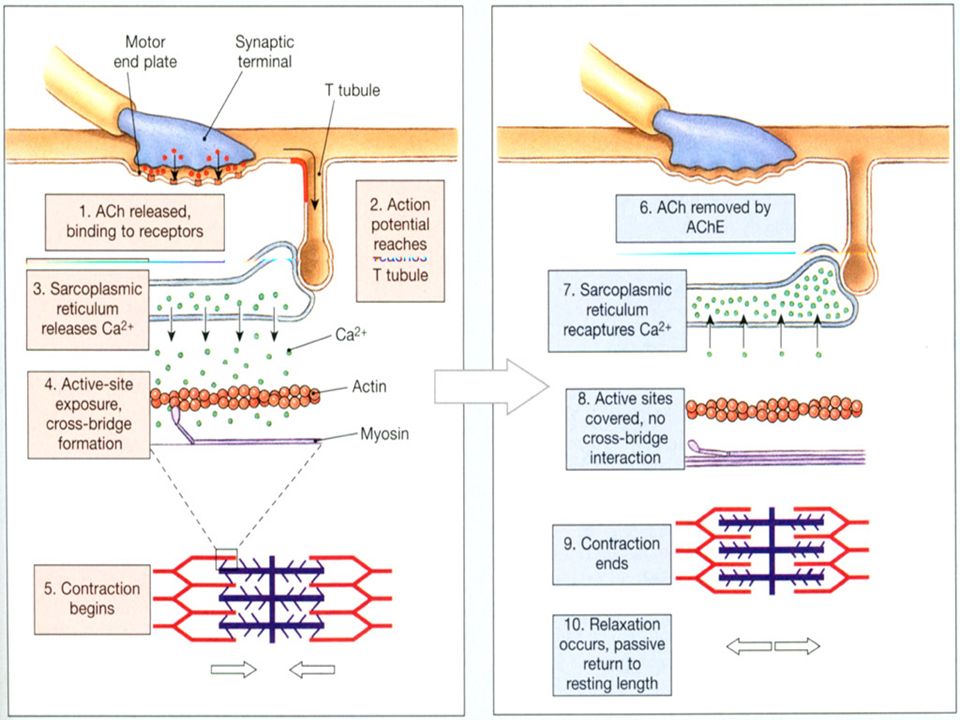
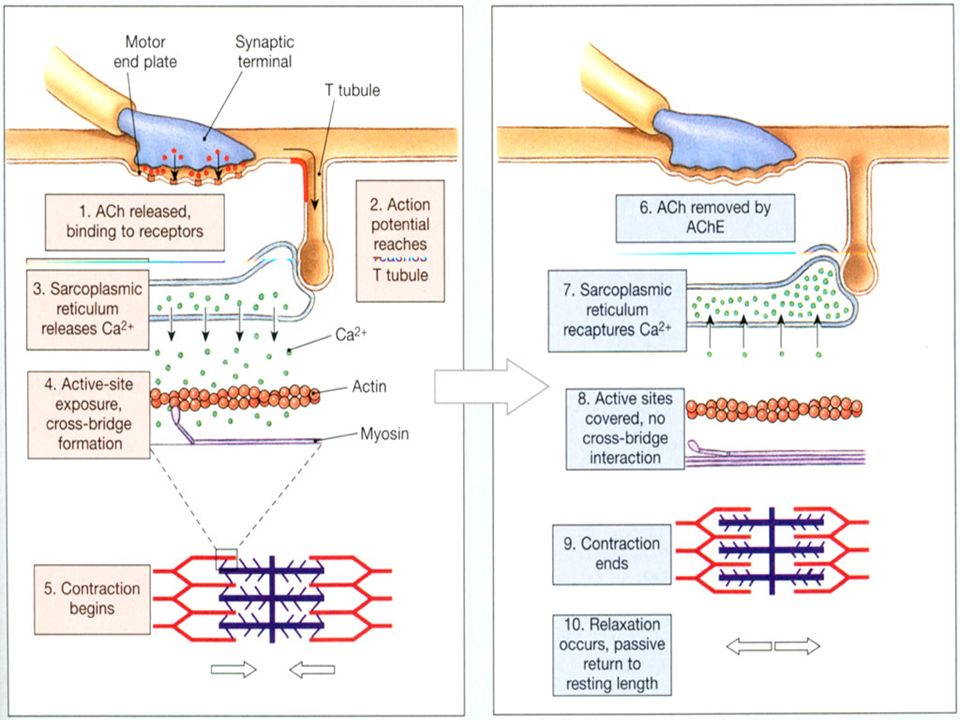
EKSITASJONS - KONTRAKSJONS KOBLING forts. 3. Ca 2+ tjener som signal til å sette i gang “crossbridge cycle” og kontraksjonen settes i gang

38
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001

39
En muskelcelle genererer kraft kontinuerlig gjennom en kontraksjon fordi “crossbridge cycle” gjennomføres simultant - men i forskjellige faser av hodebevegelse -Ca 2+ trengs for kontakt myosin-actin -ATP er energikilden som trengs for “åretak” I forbindelse med en kontraksjon gjennomfører hvert myosinhode omkring 5 cycler/sek, men fordi hvert tykt filament har flere hundre hoder, kan tusenvis av kraftutladninger utløses. Oppsummering – muskelkontraksjonen

40
En muskelcelle stopper å kontrahere når signalet fra motorneuronet opphører, og aksjonspotensialet ikke lengre er tilstede i sarkolemma. Når et aksjonspotensiale medfører Ca 2+ frigjøring, vil dette ikke kunne pågå “i det uendelige”. Når Ca 2+ konsentrasjonen stiger, vil Ca 2+ starte med å binde seg til spesielle seter i det sarcoplasmatiske retikulum - som får Ca 2+ -kanalene til å lukke seg stopper frigjøringen av Ca 2+ stimulerer aktiv transport av Ca 2+ tilbake til det sarcoplasmatiske reiculum

41
Motoriske forhornsceller

42
Motorisk enhet: enheten bestående av et motorisk neuron og de muskelfibre som innerveres. Latensperioden: tiden mellom starten på aksjonspotensialet til muskelkontraksjonen begynner Kontraksjonsfasen: perioden fra slutten av latensperioden til muskelkontraksjonen når toppen Relaksasjonsfasen: perioden mellom toppen av muskelkontraksjonen og til slutten av kontraksjonen

43
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001

45
MUSKELCELLEMETABOLISMEN OKSIDASJON Glucose + 60 2 6CO 2 + 6H 2 O (16-C) fettsyre + 23O 2 16CO 2 + 146H 2 O GLYKOLYSE Glucose 2 lactat 36ADP + P i 36ATP 131ADP + P i 131ATP 2ADP + P i 2ATP
 fettsyre + 23O 2 16CO H 2 O GLYKOLYSE Glucose 2 lactat 36ADP + P i 36ATP 131ADP + P i 131ATP 2ADP + P i 2ATP")
46
MUSKELCELLEMETABOLISME OG KONTRAKSJONENS INTENSITET/VARIGHET JEVN/MODERAT KONTRAKSJON. Mesteparten av ATP skaffes til veie ved oksydativ fosforylering. de første sekundene er musklene avhengig av lagret glykogen for å sørge for glukosetilførsel til ATP produksjonen. Deretter er energibehovet avhengig av tilført glukose, og etter 30 min blir fettsyrer den dominerende energikilden. HARD TRENING. Oksydativ fosforylering mindre betydningsfull som kilde for ATP produksjon, og substrat-nivå fosforylering (spesielt glykolyse) blir viktigere.Ett resultat av denne er produksjon av melkesyre (lactic acid), som akkumulerer i muskelcellen.
 blir viktigere.Ett resultat av denne er produksjon av melkesyre (lactic acid), som akkumulerer i muskelcellen..")
47
Mye av O 2 som tas opp forbrukes i mitokondrier i muskulatur Mye av CO 2 som dannes kommer fra mitokondrier i muskulatur Ved kraftanstrengelser vil ikke nok O 2 være tilgjengelig og da vil glykolyse i cytoplasma føre gi ATP + laktat. Laktat transporteres til lever og kan omsettes til glukose

48
Cori syklus 2 6

49
Alle muskelfibre kan produsere ATP basert på oksydativ fosforylering og substrat-nivå fosforylering, men evnen til det ene eller andre varierer. Basert på denne evnen, deles muskelfibre inn i to kategorier: GLYKOLYTISKE FIBRE. Høy konsentrasjon av glykolytiske enzymer i cytosol kan raskt generere ATP via glykolyse (substratnivå fosforylering). Har få mitochondrier liten kapasitet for oksydativ fosforylering. OKSYDATIVE FIBRE. Rik på mitokondrier høy kapasitet for ATP produksjon via oksydativ fosforylering. Lav konsentrasjon av glykolytiske enzymer. Myoglobin (“rød” muskulatur).
. Har få mitochondrier liten kapasitet for oksydativ fosforylering. OKSYDATIVE FIBRE. Rik på mitokondrier høy kapasitet for ATP produksjon via oksydativ fosforylering. Lav konsentrasjon av glykolytiske enzymer. Myoglobin ( rød muskulatur)..")
50
RASKE OKSYDATIVE FIBRE: Høy kapasitet for oksydativ fosforylering. Rask myosin. Moderat diameter moderat kraft LANGSOMME OKSIDATIVE FIBRE: Høy kapasitet for oksydativ fosforylering. Langsom myosin. Liten diameter liten kraft RASKE GLYKOLYTISKE FIBRE: Høy glykolytisk kapasitet. Rask myosin. Stor diameter stor kraft Melkesyre ved stor påkjenning Om et individ har mye raske (“lyse”) eller langsomme (“røde”) fibre avhenger dels av genetikk og dels av trening.
 eller langsomme ( røde ) fibre avhenger dels av genetikk og dels av trening..")
51
ANDRE ATP - GENERERENDE SYSTEMER ATP dannelse fra kreatinfosfat kreatinfosfat 5X ATP ATP dannelse fra ADP ved hjelp av myokinase 2 ADP ATP + AMP myokinase

52
PATOLOGISKE TILSTANDER MED UTGANGSPUNKT I MUSKULATUR Progressiv svekkelse av muskulatur (NB! mange kliniske videoer på YouTube). Duchennes muskeldystrofi. Insidens: ca. 1/3500 fødte, X-bundet (gutter blir syke) Mutasjoner i dystrofin-genet, koder for stort protein (427 kD). Dystrofin-proteinet inngår i kompleks som forankrer actin i cytoskjelettet til sarcolemma. Dystrofin uttrykkes også i hjertemuskulatur. Defekt fører til at sarcolemma revner muskelcellene degenererer. Pasientene dør unge (20-årene) oftest av respirasjonssvikt eller hjertesvikt. Becker’s muskeldystrofi. Rammer ca. 5/100.000 gutter. Mutasjoner i dystrofin-genet, men noe funksjon bevart. Lever til 40-årsalderen; noen har normal livslengde. Limb-girdle muskeldystrofi. Ikke kjønnsbundet. Heterogen samling av muskeldystrofier, 6 ulike genloci, oftest i subenheter av sarcoglycan-komplekset. relativt mildt forløp.
. Duchennes muskeldystrofi. Insidens: ca. 1/3500 fødte, X-bundet (gutter blir syke) Mutasjoner i dystrofin-genet, koder for stort protein (427 kD). Dystrofin-proteinet inngår i kompleks som forankrer actin i cytoskjelettet til sarcolemma. Dystrofin uttrykkes også i hjertemuskulatur. Defekt fører til at sarcolemma revner muskelcellene degenererer. Pasientene dør unge (20-årene) oftest av respirasjonssvikt eller hjertesvikt. Becker’s muskeldystrofi. Rammer ca. 5/ gutter. Mutasjoner i dystrofin-genet, men noe funksjon bevart. Lever til 40-årsalderen; noen har normal livslengde. Limb-girdle muskeldystrofi. Ikke kjønnsbundet. Heterogen samling av muskeldystrofier, 6 ulike genloci, oftest i subenheter av sarcoglycan-komplekset. relativt mildt forløp..")
53
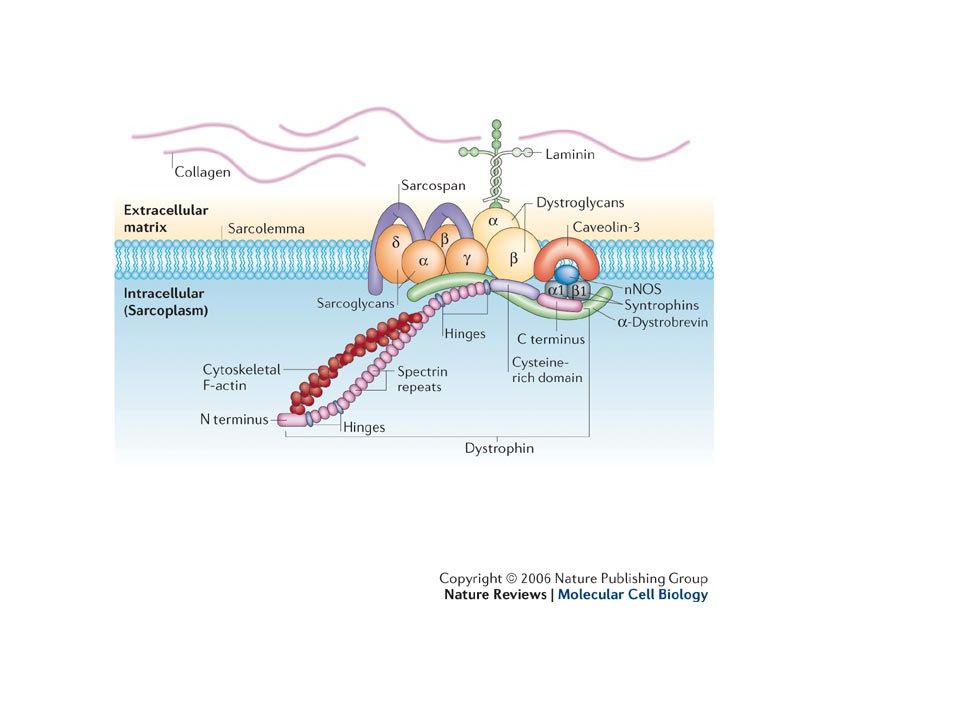
Dystrofin i kompleks med andre proteiner forankrer muskelfilamenter til sarcolemma

55
GLATT MUSKULATUR har tykke og tynne filamenter som er arrangert i parallell, men også på skrå kontraksjoner vil gå langs flere akser muskelfibrene er ikke arrangert i sarcomerer stripene mangler dense bodies, knutepunkter mellom muskel- filamenter og bindevev inne i cellen, bidrar til å overføre kontraksjonen til cellens ytre

56
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001

57
KONTRAKSJON AV GLATT MUSKULATUR er også regulert av intracellulært Ca 2+ en stor del av Ca 2+ kommer fra utsiden av cellen. Depolarisering åpning av spenningsavhengige Ca 2+ kanaler i plasmamembranen Ca 2+ vandrer intracellulært (depolarisering induserer også frigjøring av Ca 2+ fra det endoplasmatiske reticulum) Ca 2+ aktiverer “crossbridge cycle”. Ca 2+ binder til calmodulin konfigurasjonsendring aktiverer myosin kinase katalyserer fosforylering av myosinbroene aktivering (resten av cyclus tilsvarer den vi finner ved tverrstripet muskulatur).
 Ca 2+ aktiverer crossbridge cycle . Ca 2+ binder til calmodulin konfigurasjonsendring aktiverer myosin kinase katalyserer fosforylering av myosinbroene aktivering (resten av cyclus tilsvarer den vi finner ved tverrstripet muskulatur)..")
58
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001

59
HJERTEMUSKULATUR er stripete, har sarkomer-struktur kontraksjoner er regulert av troponin/ tropomyosin systemet mange gap-junctions, sørger for at aksjonspotensialet overføres i hele nettverket

60
HJERTEMUSKULATUR forts. Andre kjennetegn: LANGVARIG AKSJONSPOTENSIAL (varer omtrent like lenge som det tar hjertemuskulaturen å kontrahere og å slappe av) innebærer at summasjon av kontraksjoner ikke lar seg gjøre. Nettverket fungerer som en enhet. SINUS- OG ATROVENTRIKULÆR KNUTER. Har pacemaker-aktivitet. Det innebærer at aksjonspotensialet i hjertemuskulatur ikke avhenger av neural stimulering (myogen kontraktil aktivitet).
 innebærer at summasjon av kontraksjoner ikke lar seg gjøre. Nettverket fungerer som en enhet. SINUS- OG ATROVENTRIKULÆR KNUTER. Har pacemaker-aktivitet. Det innebærer at aksjonspotensialet i hjertemuskulatur ikke avhenger av neural stimulering (myogen kontraktil aktivitet)..")
61
Germann WJ, Stanfield CL. Pronciples of human physiology. Benjamin Cummings 2001 OPPSUMMERING

62
hele energimetabolismen i muskelceller er rettet mot høy ATP produksjon i tillegg til “vanlige” synteseveier har muskler spesielle ATP genererende systemer

63
OPPSUMMERING VIKTIGSTE ENERGIKILDER: oksydasjon av glukose og fettsyrer (aerobt, lett arbeid) glykolyse (anaerobt, tungt arbeid)
 glykolyse (anaerobt, tungt arbeid)")
64
OPPSUMMERING EFFEKTIVITET OKSYDASJON Glucose + 60 2 6CO 2 + 6H 2 O (16-C) fettsyre + 23O 2 16CO 2 + 146H 2 O GLYKOLYSE Glucose 2lactat 36ADP + P i 36ATP 131ADP + P i 131ATP 2DP + P i 2ATP
 fettsyre + 23O 2 16CO H 2 O GLYKOLYSE Glucose 2lactat 36ADP + P i 36ATP 131ADP + P i 131ATP 2DP + P i 2ATP")
65
OPPSUMMERING Andre ATP genererende systemer: ATP dannelse fra kreatinfosfat (kreatinfosfatkons. = 5X ATP kons.) kreatinkinase kreatinfosfat + ADP ATP + kreatin ATP dannelse fra ADP ved hjelp av myokinase 2 ADP ATP + AMP myokinase
 kreatinkinase kreatinfosfat + ADP ATP + kreatin ATP dannelse fra ADP ved hjelp av myokinase 2 ADP ATP + AMP myokinase.")
66
Boron WF, Boulpaep EL. Medical physiology. Saunders 2003

Liknende presentasjoner