Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
Regulering av karbohydratmetabolismen
Winnie Eskild. IMBV 2004

2
Glykogen glykogenese glykogenolyse Glukose-6-fosfat glukoneogenese glykolyse Pyruvat

3
Nedbrytning av glykogen

4
Glykogen nedbrytning

5
Regulering av glykogenkatabolisme
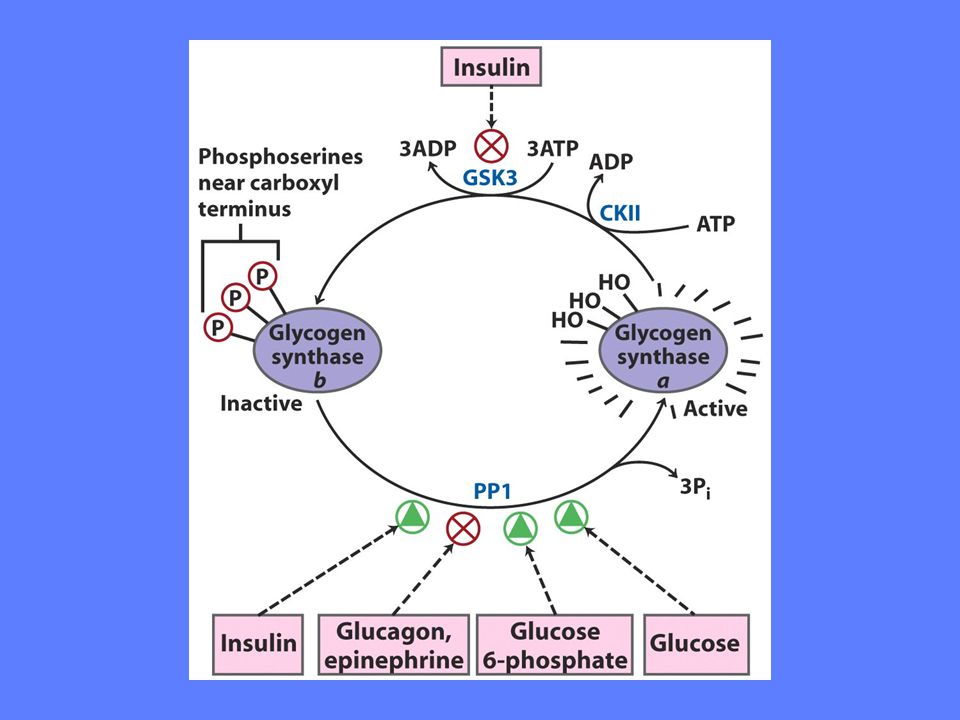
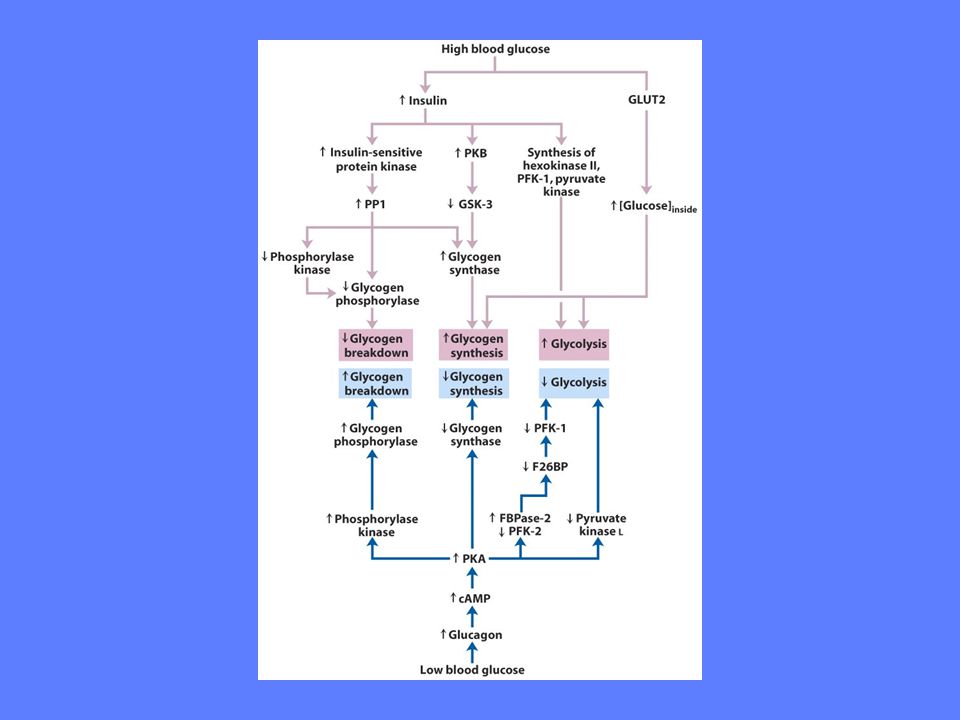
Glykogen fosforylase stimuleres av hormonene: glukagon, adrenalin Glykogen fosforylase reguleres allosterisk av glukose Glykogen fosforylase hemmes av hormonet: insulin

6
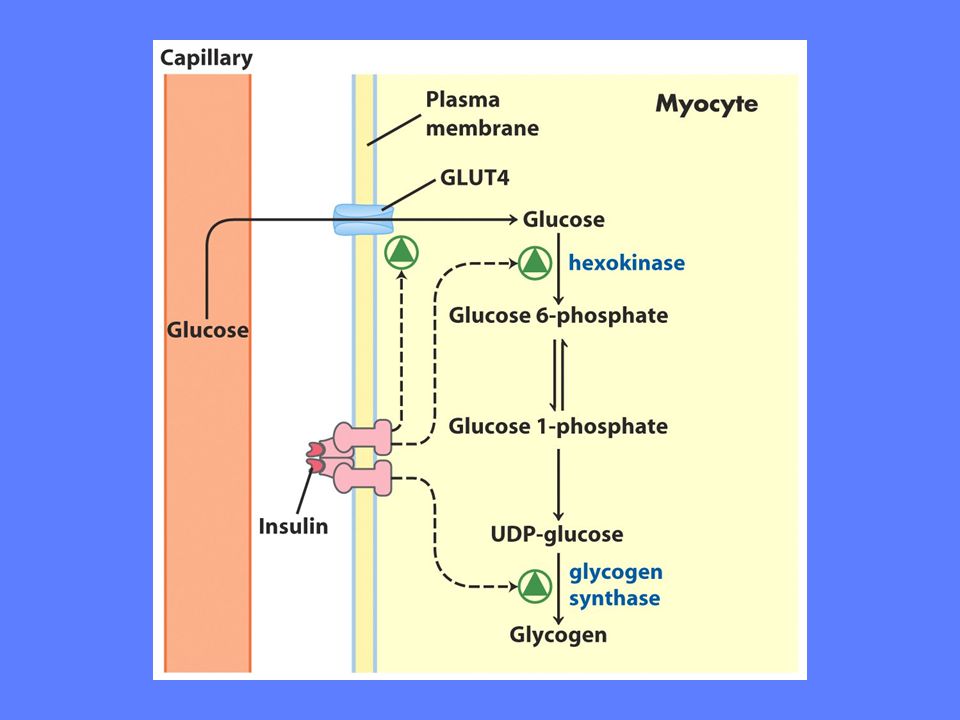
Glykogensyntese Ved overskudd av glukose dannes lagringsformer som varierer avhengig av organisme og celletype Dyr (vertebrater) og mikroorganismer lager glykogen, mens planter lager stivelse Alle dyrevev kan lage glykogen men i praksis er det bare lever og muskel som gjør det Leverens glykogen benyttes til å opprettholde riktig konsentrasjon av glukose i blodet Musklenes glykogen er et glukoselager som kan brukes ved intens arbeid Glukose eller andre heksoser må aktiveres før de kan polymeriseres Aktiveringen består av påkopling av nukleotider via en fosfoester-binding For glykogensyntese trengs det UDP-glukose (UDP = uridindifosfat)
 og mikroorganismer lager glykogen, mens planter lager stivelse. Alle dyrevev kan lage glykogen men i praksis er det bare lever og muskel som gjør det. Leverens glykogen benyttes til å opprettholde riktig konsentrasjon av glukose i blodet. Musklenes glykogen er et glukoselager som kan brukes ved intens arbeid. Glukose eller andre heksoser må aktiveres før de kan polymeriseres. Aktiveringen består av påkopling av nukleotider via en fosfoester-binding. For glykogensyntese trengs det UDP-glukose (UDP = uridindifosfat)")
7
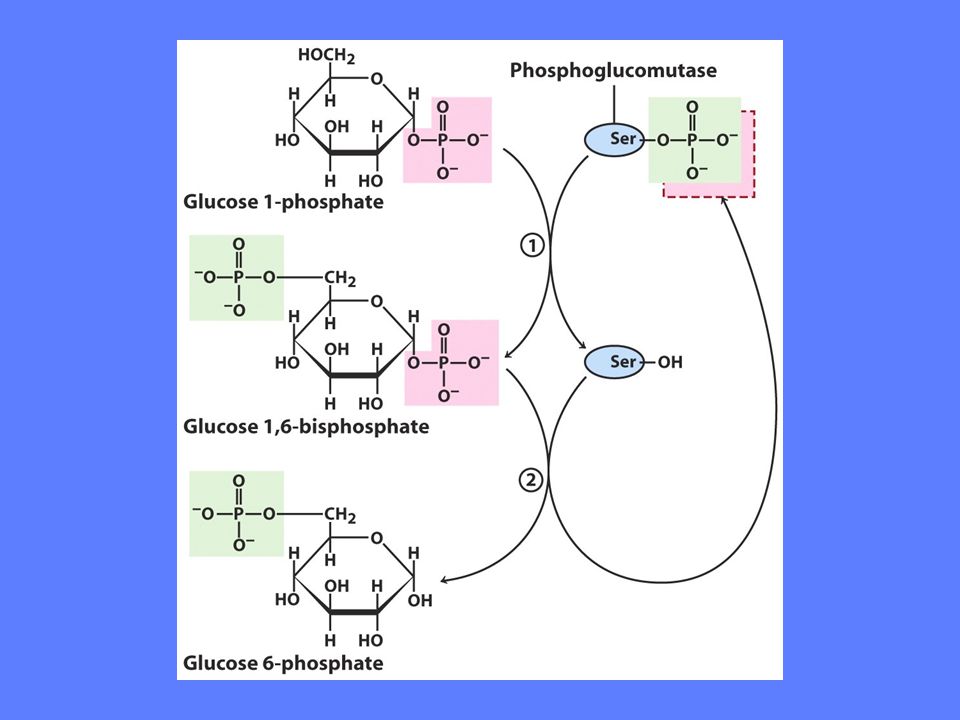
Glukose til glukose-1-fosfat
Omdannelsen av glukose til glukose-1-fosfat krever to enzymer Enzymet er heksokinase (lever) eller glukokinase (muskel) Glukose + ATP => glukose-6-fosfat Enzymet er fosfoglukomutase glukose-6-fosfat <=> glukose-1-fosfat Mye av den glukose som går inn i disse reaksjoner kommer fra glukoneogenesen som har omdannet laktat fra røde blodceller til glc-6-P
 eller glukokinase (muskel) Glukose + ATP => glukose-6-fosfat. Enzymet er fosfoglukomutase. glukose-6-fosfat <=> glukose-1-fosfat. Mye av den glukose som går inn i disse reaksjoner kommer fra glukoneogenesen som har omdannet laktat fra røde blodceller til glc-6-P.")
9
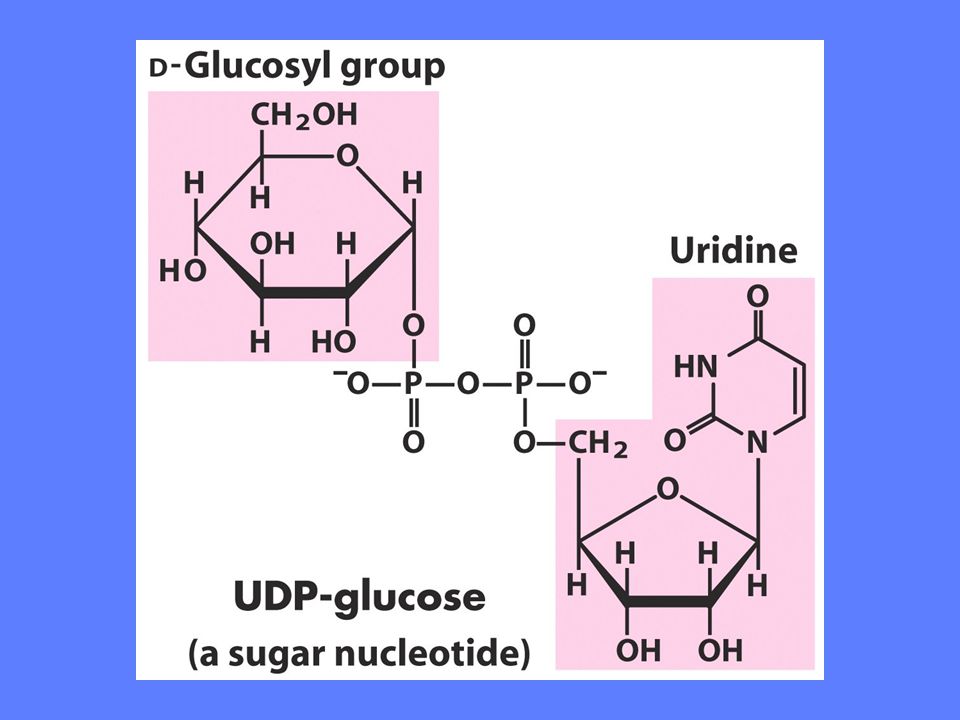
UDP-glukose pyrofosforylase + pyrofosfatase
G’o = -20 kJ/mol

11
Sukker-nukleotidenes fortrinn
Reaksjonen er irreversibel i cellen. Nesten all den Gibbs frie energi som frigis ved hydrolyse av PPi brukes til å drive reaksjonen Sukker-nukleotidets størrelse gir gode muligheter for frigjøring av bindingsenergi ved dannelse av enzym-substrat komplekser. Dette bidrar til økt spesifisitet og økt reaksjonshastighet Koplingen av nukleotidet til C-atom 1 i heksosen gjør denne til et attraktivt mål for et nuklefilt angrep under katalysen Ved å kople et nukleotid på heksosen gjøres den utilgjengelig for en del andre reaksjoner. Den reserveres for glykogensyntese

12
1=>4 glykosidbinding
Glykogensyntese Glykogen syntase kopler C-atom 1 fra innkommende glukose til C-atom 4 i eksisterende glykogen. C1 er det anomere C-atom og reduserende ende. C4 er den ikke reduserende enden 1=>4 glykosidbinding Glykogensyntase kan ikke lage 1=>6 bindinger og må ha en primer på minst 8 glukoseenheter koplet med 1=>4 bindinger

13
Glykogen branching-enzyme lager forgreninger
Amylo (1=>4) to (1=>6) transglykosylase/ glykosyl-(4=>6)-transferase flytter de siste 6-7 glukoseenheter i en glukosepolymer til C-atom 6 på en glukoseenhet lenger inne i kjeden Forgrening øker antall ikke-reduserende ender (syntese/degradering) og løselighet
 to (1=>6) transglykosylase/ glykosyl-(4=>6)-transferase flytter de siste 6-7 glukoseenheter i en glukosepolymer til C-atom 6 på en glukoseenhet lenger inne i kjeden. Forgrening øker antall ikke-reduserende ender (syntese/degradering) og løselighet.")
14
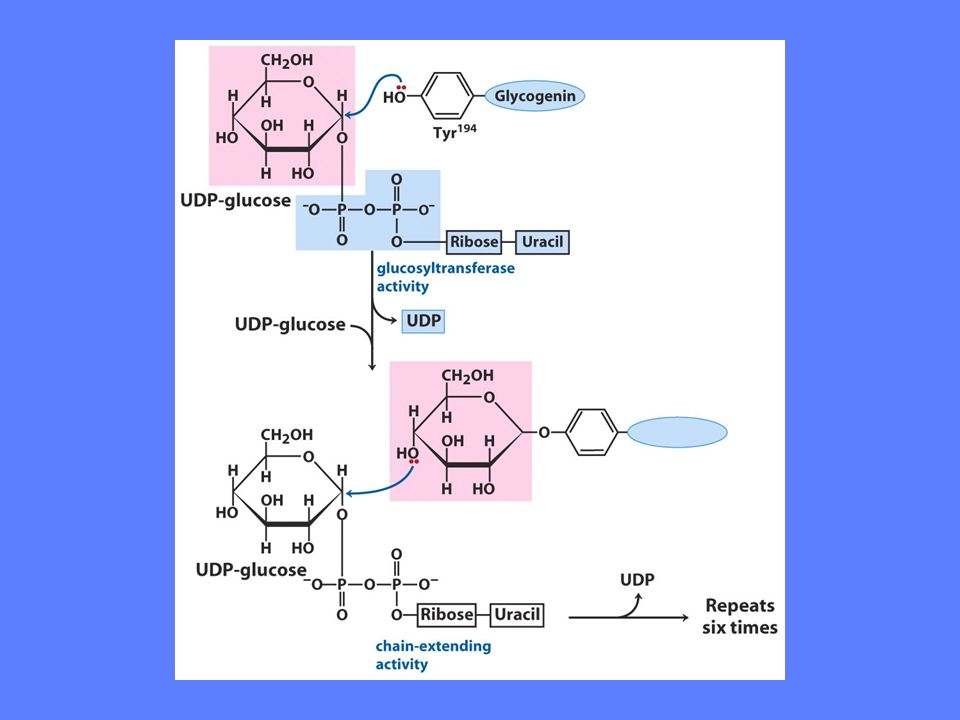
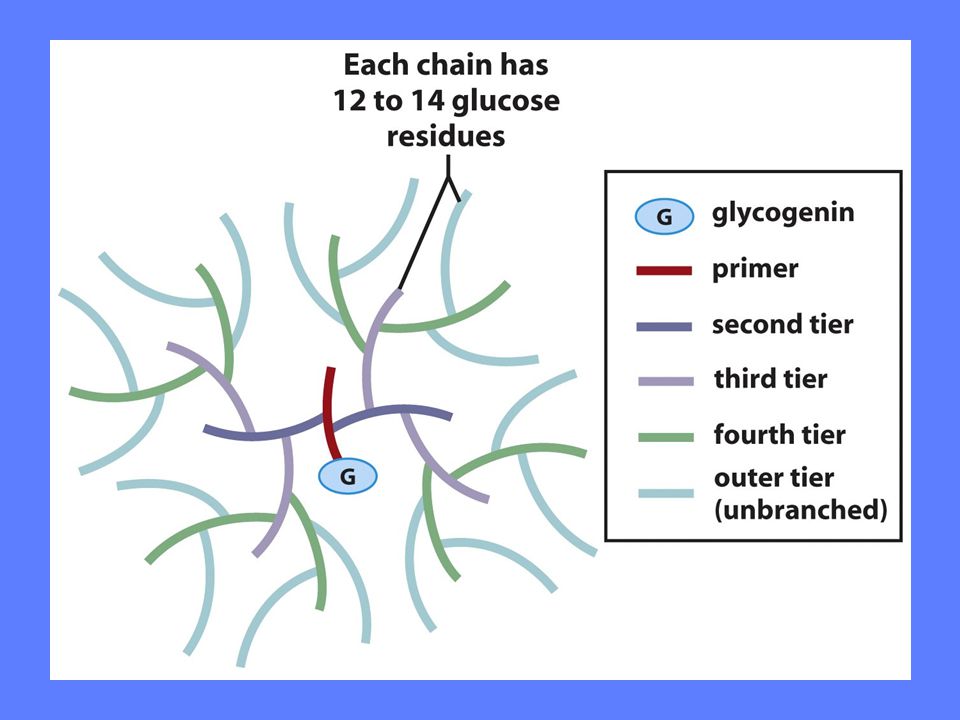
Hvordan lages nye glykogenmolekyler
Glykogenin fungerer som primer og enzym ved syntese av nye glykogenmolekyler Tyr194 glykosyleres av glykosyltransferase UDP-glukose leverer glukosemolekylet Glykogen syntase koples på Glykosyltransferase bygger opp glukosepolymeren til ca 7 glukoseenheter Glykogen syntase overtar polymeriseringen og polymeren frigjøres etterhvert I sentrum av hvert glykogenkorn ligger et glykogenin bundet til en redusernde ende

17
Dimer av glukogenin med UDP-glukose

18
Regulering av glykogen syntesen
Glykogen syntase stimuleres av insulin, glukose, glukose-6-fosfat Glykogen syntase hemmes av glukagon, adrenalin

19
Regulering av glukogenmetabolismen
Glykogen + + Glukagon Adrenalin Kalsium Glukose Glc-6-P Insulin - - Glukagon Adrenalin Insulin Glukose UDP-Glukose/Glukose-1-fosfat

21
Nedbrytning av glykogen

22
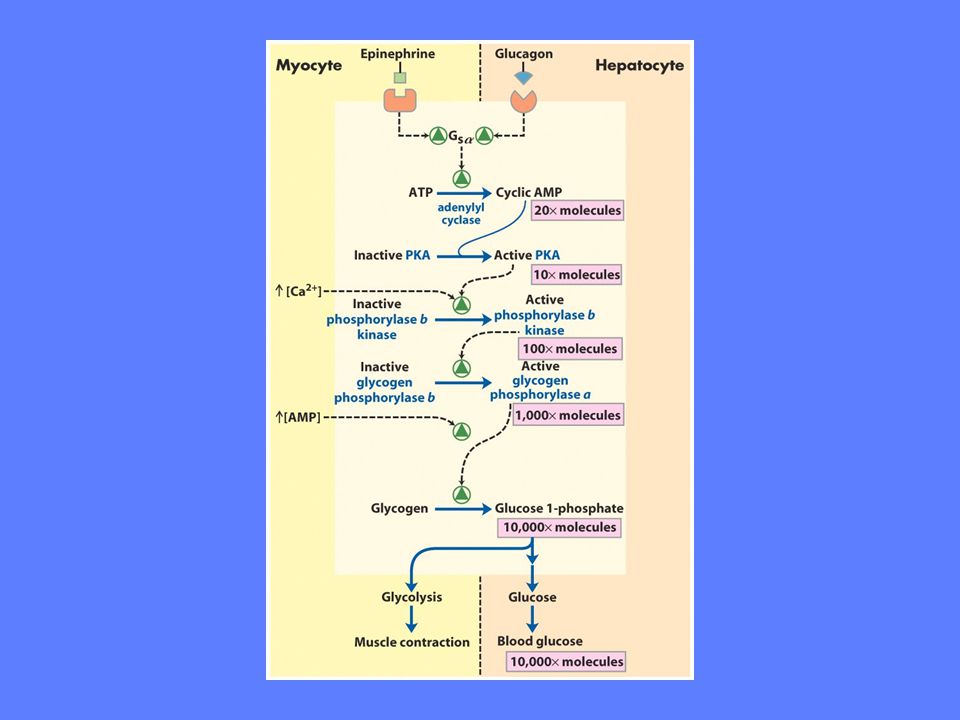
Forskjellige hormoner aktiverer fosforylase i lever og muskel
Glukagon øker ved lav blodglukose Muskel: Adrenalin og kalsium øker ved muskel- aktivitet

23
Binding av glukose medfører konformasjonsendring
Binding av glukose medfører konformasjonsendring. To fosforylerte serinsidekjeder blir tilgjengelig.

24
Enzymet fosforylase a fosfatase defosforylerer fosforylase a.
Resultatet er den mindre aktive fosforylase b

26
Hastighetsbegrensende reaksjoner
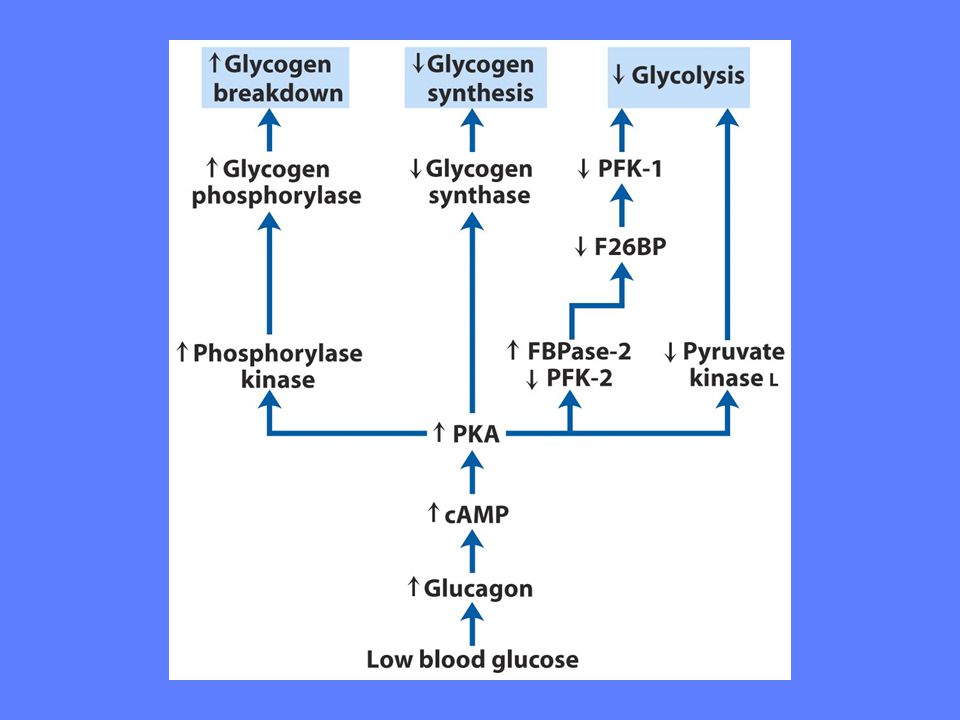
Rate-limiting steps Disse enzymkontrollerte reaksjoner finnes hvor et substrat kan ta flere veier Cellen er avhengig av å ha en konstant nivå av ATP hele tiden Cellens behov varierer fra minutt til minutt og alle enzymkontrollerte reaksjonsveier er avstemt etter behovene

27
Regulering av glykolyse og glukoneogenese

28
Regulering av glykolysen
Tre viktige reguleringspunkter: heksokinase, fosfofruktokinase-1 og pyruvat kinase Dannelse av produkt i de fleste enzymkatalyserte reaksjoner er bestemt av tilgangen på substrat Dette gjelder ikke disse regulerte enzymer som er allosterisk regulert Her bestemmer andre forhold hvor mye produkt som skal dannes uansett tilgang på substrat

29
Regulering av heksokinase
Heksokinase er allosterisk regulert (hemmet) av sitt eget produkt Heksokinasen i muskel har lav Km (0,1 mM). Dette enzymet arbeider i takt med tilgang på substrat hele tiden Heksokinase isoenzymet i lever heter glukokinase. Dette enzymet har høy Km (10 mM), dvs det arbeider bare i høy hastighet ved høye nivåer av glukose i blodet Normale blodglukosenivåer er ca 4-5 mM Glukokinase hemmes av fruktose-6-fosfat
 av sitt eget produkt. Heksokinasen i muskel har lav Km (0,1 mM). Dette enzymet arbeider i takt med tilgang på substrat hele tiden. Heksokinase isoenzymet i lever heter glukokinase. Dette enzymet har høy Km (10 mM), dvs det arbeider bare i høy hastighet ved høye nivåer av glukose i blodet. Normale blodglukosenivåer er ca 4-5 mM. Glukokinase hemmes av fruktose-6-fosfat.")
30
Heksokinase i lever hemmes allosterisk av fruktose-6-fosfat,
som får et reguleringsprotein til å binde enzymet og overføre det til cellekjernen. Heksokinase fjernes fra multienzymkomplekset av glykolyse- enzymer

32
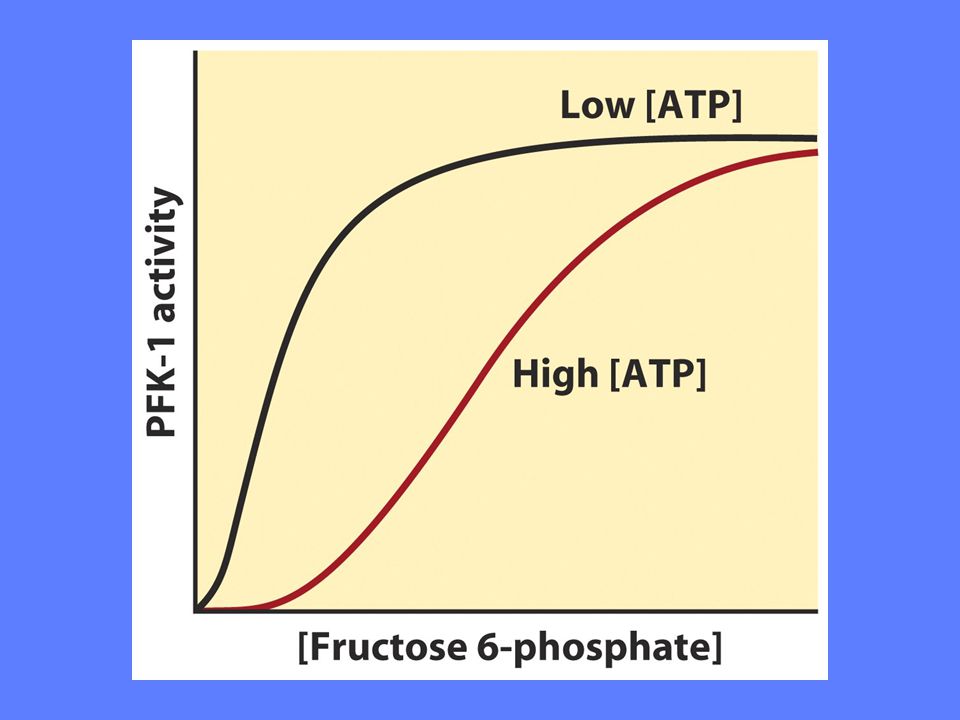
Regulering av fosfofruktokinase 1
Allosterisk regulering endrer enzymets affinitet for substrat Høyt nivå av ATP senker affiniteten for substratet. ADP og AMP hemmer effekten av ATP Høyt nivå av citrat øker ATPs hemmende effekt på enzymet

33
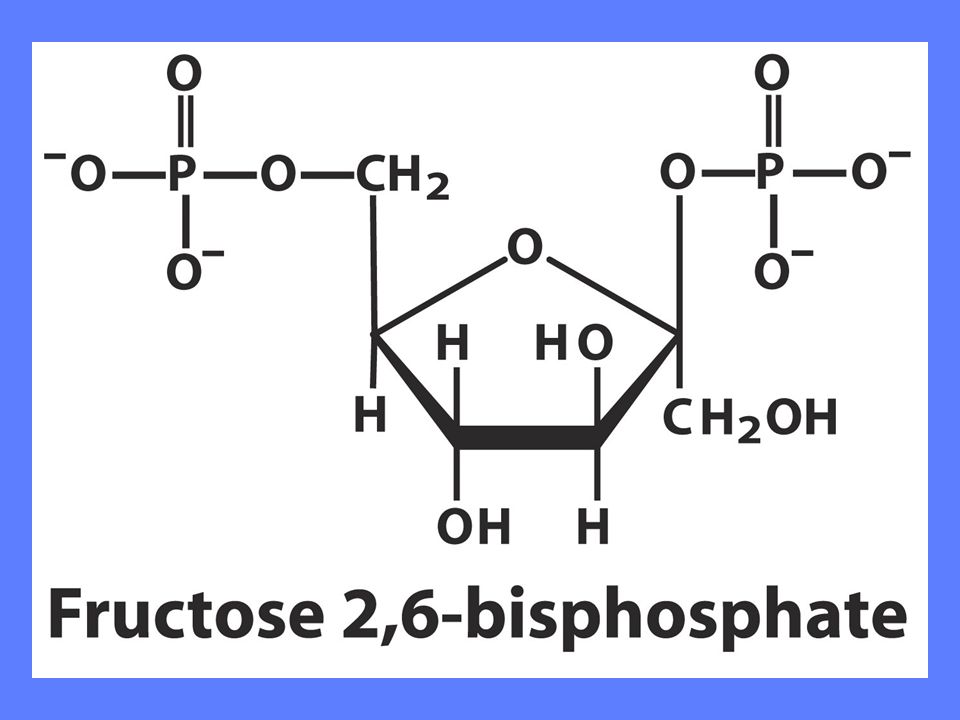
Regulering av fosfofruktokinase 1
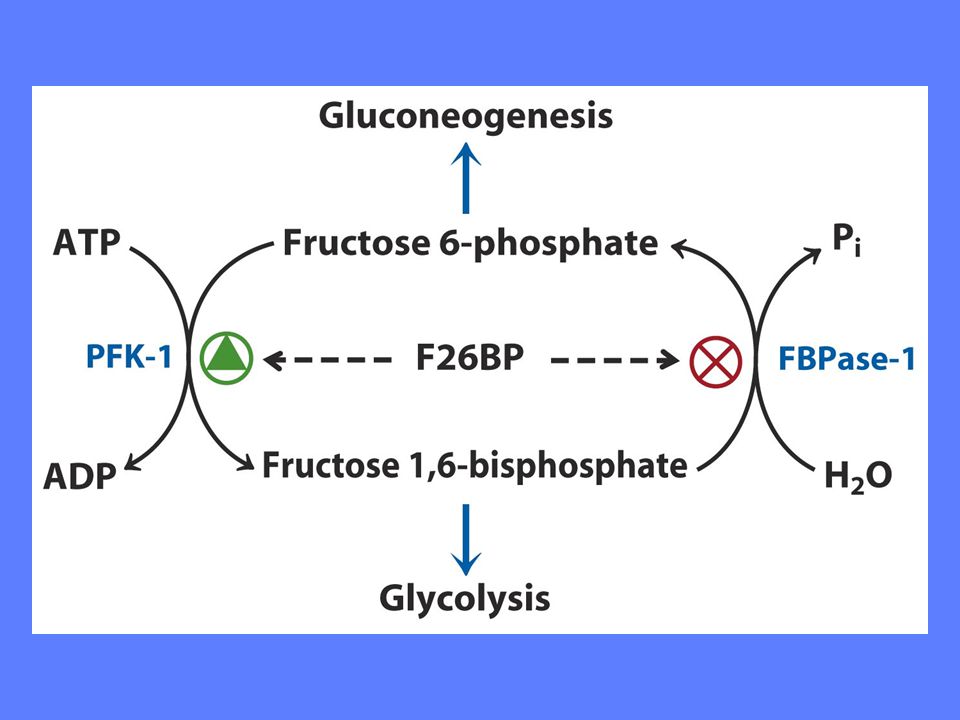
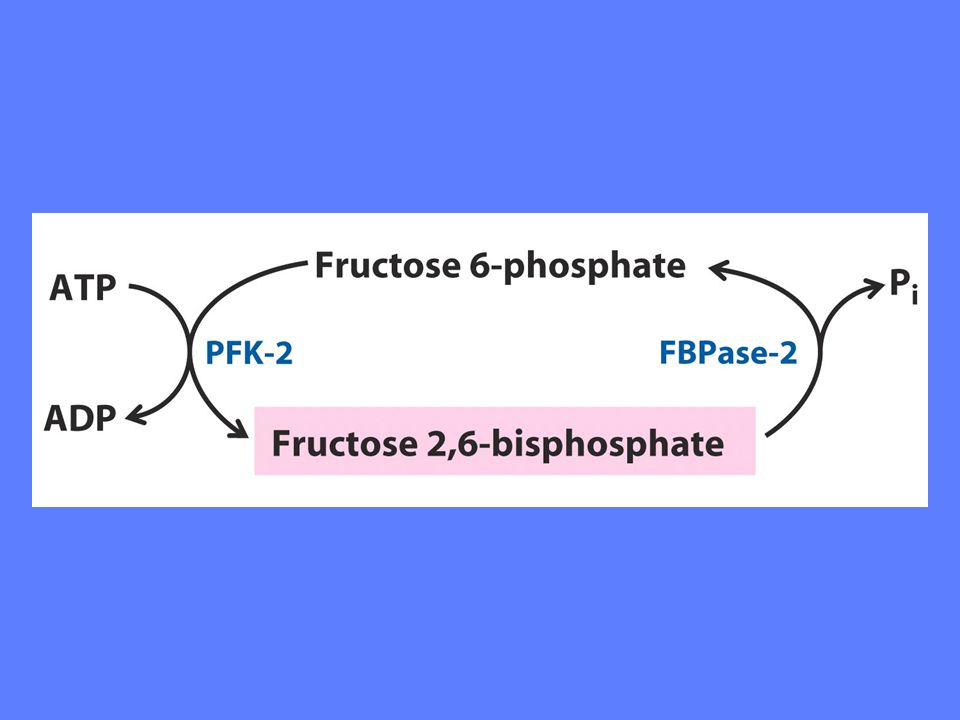
Fruktose-2,6-bisfosfat stimulerer PFK-1 aktivitet ved å øke enzymets affinitet for substrat F2BP viktigste regulator under Normale forhold Insulin øker F2BP, mens glukagon reduserer F2BP

35
Regulering av pyruvat kinase
Det finnes 3 isoenzymer som har vevsspesifikk ekspresjon Høye nivåer av ATP, acetyl-CoA og langkjedete fettsyrer hemmer aktiviteten allosterisk ved å øke K 0,5. Gjelder alle tre isoformer. I lever fører økt glukagon til inaktivering av enzymet ved fosforylering

36
Regulering av glukoneogenesen
Glukoneogesen bruker mere energi til å bygge et molekyle glukose enn cellen kan ekstrahere fra samme glukosemolekyle når det brytes ned til pyruvat Derfor er det svært viktig at disse to motsatt rettede prosesser ikke foregår samtidig Det er to viktige reguleringspunkter: Pyruvat karboksylase og fruktose 1,6-bisfosfatase.I tillegg er pyruvat karboksylase regulert

37
Regulering av pyruvat karboksylase
Høye nivåer av acetyl-CoA viser at cellen har tilstrekkelig substrat til energiformål. Dette stimulerer pyruvat karboksylase og dirigerer dermed pyruvat til gluko-neogenese. Lave nivåer av acetyl-CoA stimulerer pyruvat dehydrogenase som dirigerer pyruvat til nedbrytning via sitronsyre syklusen

38
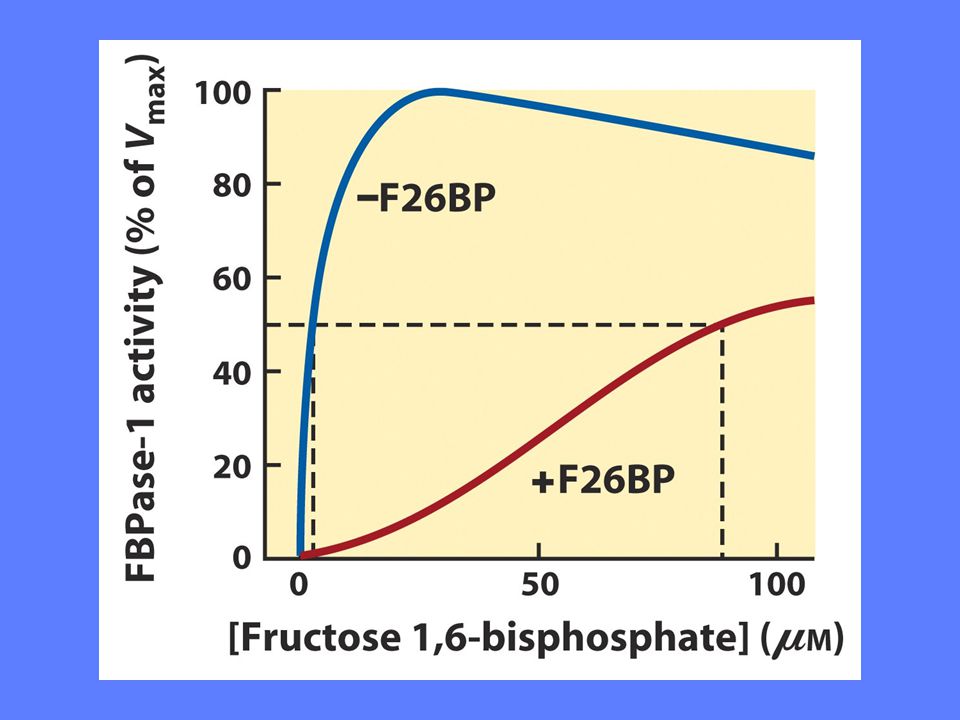
Regulering av fruktose-1,6-bisfosfatase
AMP og fruktose-2,6-bisfosfat hemmer dette enzymet

42
Glukose-6-fosfatase finnes bare i
leverens endoplasmatisk retikulum Denne lokalisering hindrer at glucose-6-fosfat som skal gjennom glykolysen slipper ut av cellen

43
”Futile cycles” Fruktose-6-P + ATP => fruktose 1,6-bisfosfat + ADP
Fruktose 1,6-bisfosfat + H2O => fruktose-6-P + Pi Sum: ATP + H2O => ADP + Pi + varme Ved å la disse to motsattrettede reasjoner gå samtidig produseres varme og intet annet Denne ”syklus” gir også mulighet til å kontrollere hvilken retning substratet skal ta. Her kan glykolyse balanseres mot glukoneogenese I visse situasjoner er det ønskelig som f. eks når humlen skal varme opp vingene sine til 30oC før flyveturen på en kjølig dag ”Futile cycles” har samme funksjon som avkopling av oksidativ fosforylering i brunt fettvev

Liknende presentasjoner





















>")