Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
Bioenergetiske prinsipper
Forutsetningen for at biokjemiske prosesser forløper Winnie Eskild, IMBV 2004

2
Metabolisme = katabolisme + anabolisme

3
Katabole prosesser konvergerer Anabole prosesser divergerer

4
Hva driver biokjemiske reaksjoner
Forskell mellom Gibbs fri energi i startprodukt og sluttprodukt Ved de rette forholdene vil en biokjemisk reaksjon forløpe til likevekt oppnås Ved likevekt går reaksjonen like fort i begge retninger aA + bB <=> cC + dD Likevektskonstanten er da Keq= C c D d A a B b DG’o er definert for fysiologiske forhold hvor pH er 7, H2O er 55,5 M og Mg 2+ er 1 mM. Gjelder også K’eq Drivkraften i en reaksjon , DG’o og K’eq har følgende relation DG’o = -RTln K’eq

5
Sammenheng mellom DG’o og K’eq Eksponentielt forhold mellom DG’o og K’eq: Små endringer i DG’o => store endringer i K’eq Bare litt negativ DG’o => likevekten sterkt forskjøvet mot produkt

6
Sammenheng mellom DG’o og K’eq

7
Glukose-1-fosfat og glukose-6-fosfat
Når enzym er tilstede vil det ved likevekt være 19 ganger mere glukose-6-fosfat uansett om utgangspunktet er 1 M glukose-1-fosfat eller 1 M glukose-6-fosfat. Enzyme: fosfoglukomutase. Glukose-1-fosfat <=> glukose-6-fosfat K’eq = glukose-6-fosfat = 0,95 M = 19 glukose-1-fosfat ,05 M Ut fra denne likevektskonstant kan vi beregne endring i standard fri energi DG’o = -RT ln Keq = -(8,315 J/mol x K)(298 K)(ln 19) = J/mol = - 7,3 kJ/mol
(298 K)(ln 19) = J/mol. = - 7,3 kJ/mol.")
8
I cellen finnes ingen standard konsentrasjoner
DG = DG’o + RT ln C D A B Her er A,B,C og D de aktuelle konsentrasjonene Ved likevekt er DG = 0 Vi får da at DG’o = -RT lnK’eq Det er DG som avgjør om en reaksjon forløper, selvom DG’0 er positiv kan godt DG være negativ Ved å fjerne produktene så snart de er dannet holdes K’eq under 1 og stor nok til at hele uttrykket blir negativt

9
DG’o verdier kan adderes
Glukose + Pi Glukose-6-fosfat + H2O DG’o = 13,8 kJ/mol ATP + H2O ADP + Pi DG’o = -30,5 kJ/mol Glukose + ATP Glukose-6-fosfat + ADP DG’o = -16,7 kJ/mol De to delreaksjoner har hver sin DG’o men det er sumreaksjonen som teller i biokjemien Derved blir en endergon reaksjon mulig ved kopling til en eksergon reaksjon slik at den totale reaksjonen er eksergon

10
ATP som energileverandør
Når ATP deltar i reaksjoner og etter endt reaksjon foreligger som ADP + Pi eller AMP + 2 Pi sir vi at hydrolysering av ATP har levert energi til å drive reaksjonen I de fleste tilfelle forekommer en gruppeoverføring fra ATP til et mellomprodukt. Denne gruppen kan være fosfat, pyrofosfat eller AMP

11
Overføring av aminogruppe til glutamat koster ATP

12
Aktivering av en fettsyre koster ”2” ATP

13
Grunnlaget for det høye energinivået i bindingene mellom fosfatgruppene i ATP
Figur 14-1

14
ATP ATP har 3 negativ ladete fosfatgrupper: elektrostatisk frastøting
Fraspalting av en fosfatgruppe gir mulighet for resonansstabilisering og den gjenværende ADP kan dissosiere og frigi en H+ til omgivelsene hvor konsentrasjonen av protoner er svært lav ATP 4- + H2O ADP 3- + Pi 2- + H+ DG’o = - 30,5 kJ/mol Denne gjelder under standardbetingelser. I cellen er ATP tilstede i helt andre konsentrasjoner og i tillegg kompleksbundet med Mg 2+ I cellen vil en derfor oftere se DG på kJ/mol

15
ATP Hydrolyse av a-b anhydrid-bindingen frigir ca 46 kJ/mol
I tillegg frigis det ca 19 kJ/mol ved hydrolyse av PPi Hydrolyse av b-g anhydrid-bindingen frigir ca 31 kJ/mol

16
ATP i røde blodceller Her er konsentrasjonene: ATP: 2,25 mM, ADP: 0,25 mM, Pi: 1,65 mM DGp brukes til å angi den reelle hydrolyseringsenergi DGp = DG’o + RTln Settes verdiene inn får en: DGp = - 51,8 kJ/mol DGp kalles fosforyleringspotensialet og angir den aktuelle fri hydrolyseringsenergi i et gitt miljø DGp vil variere fra celle til celle og innen cellen avhengig av tilgang på energi, pH, andre reaktanter o.l. Total ATP, ADP og Pi i cellen kan være langt høyere enn de konsentrasjoner som er tilgjengelige for disse reaksjoner, ved f.eks. å være tett knyttet til proteiner ADP Pi ATP

17
Nukleotidnivå i forskjellige celler
Tabell 14-5

18
Andre energirike forbindelser Fosfoenolpyruvat
Fosfoenolpyruvat har energirik fosfoester binding Fosfoenolpyruvat har bare en resonansform mens pyruvat har både enolformen og ketoformen PEP 3+ + H2O pyruvat + Pi DG’o = - 61,9 kJ/mol

19
1,3-Bisfosfoglyserat 1,3-bisfosfoglyserat har en anhydridbinding med bare én resonansform Etter hydrolyse kan 3-fosfo-glyserat fradissosiere en H+ til et med miljø med lave konsentrasjoner av H+ og får i tillegg to resonansformer 1,3-bisfosfoglyserat 4- + H2O => 3-fosfoglyserat 3- + Pi 2+ + H+ DG’o = - 49,3 kJ/mol

20
Flere fosfoforbindelser

21
ATP er cellens energivaluta
ATP’s fosforyleringspotensiale er passe stort til at gjøre det svært velegnet til å ta imot og gi fra seg energi i cellens forskjellige reaksjoner ATP er stabilt fordi aktiverings-energien for hydrolyse er høy ATP må bindes til spesifikke enzymer som katalyserer prosesser hvor ATP er ”energi-leverandør” før hydrolyse er mulig Enzymene muliggjør nøye kontroll av energiforbruket

22
Thioestre frigir mye energi ved hydrolyse

23
Acetyl-coenzym A Acetyl-CoA + H2O acetat- + CoA + H+
DG’o = - 32,2 kJ/mol

24
Fire forhold som gir store, negative DG’o ved hydrolyse
1) Fjerning/reduksjon av elektrisk frastøting mellom grupper Eks: ATP 2) Stabilisering av produktene ved ionisering Eks: ATP, acylfosfater, thioestre 3) Stabilisering av produktene ved tautomerisering Eks: fosfoenolpyruvat 4) Resonanstabilisering av produktene acylfosfater, thioestre, Pi
 Fjerning/reduksjon av elektrisk frastøting mellom grupper. Eks: ATP. 2) Stabilisering av produktene ved ionisering. Eks: ATP, acylfosfater, thioestre. 3) Stabilisering av produktene ved tautomerisering. Eks: fosfoenolpyruvat. 4) Resonanstabilisering av produktene. acylfosfater, thioestre, Pi.")
25
Bioenergetiske prinsipper Ib

26
Oppgave 1 Beregn Keq ved 37oC for reaksjonen:
Fosfoarginin + ADP arginin + ATP Likevektskonsentrasjonenen av substrater og produkter er: Fosfoarginin 0,737 mM ADP ,750 mM Arginin ,78 mM ATP ,87 mM Hva er DG’o for reaksjonen?. Er den spontan under standard forhold?

27
Svar på oppgave 1 DG’o = -RT ln Keq
DG’o = -(8,3145 x 10-3 kJ x K-1 x mol-1)(298K) x ln33,5 DG’o = -8,7 kJ/mol Reaksjonen vil gå spontant under standard betingelser arginin ATP fosfoarginin ADP (4,78 x 10-3) (3,87 x 10-3) (0,737 x 10-3) (0,75 x 10-3)
(298K) x ln33,5. DG’o = -8,7 kJ/mol. Reaksjonen vil gå spontant under standard betingelser. arginin. ATP. fosfoarginin. ADP. (4,78 x 10-3) (3,87 x 10-3) (0,737 x 10-3) (0,75 x 10-3)")
28
Oppgave 2 Likevektskonstanten for omdannelse av glukose-6-fosfat til fruktose-6-fosfat er 0,41. Reaksjonen er reversibel og katalyseres av enzymet fosfoheksose isomerase. Hva er DG’o for denne reaksjonen? Hvilken retning vil reaksjonen gå under standard forhold? Hva er DG for reaksjonen ved 37oC når konsentrasjonen av glukose-6-fosfat er 2,0 mM og konsentrasjonen av fruktose-6-fosfat er 0,5 mM? Hvilken retning vil reaksjonen gå under disse forholdene?

29
Svar på oppgave 2 = -(8,3145 x 10-3 kJ/K/mol)(298K)ln 0,41
DG’o = -RT ln Keq = -(8,3145 x 10-3 kJ/K/mol)(298K)ln 0,41 = 2,2 kJ/mol Reaksjonen går mot dannelse av glukose-6-fosfat DG = DG’o + RT ln = 2,2 kJ/mol + (8,3145 x 10-4 kJ/K/mol)(310K)ln = 2,2 kJ/mol - 3,57 kJ/mol = - 1,37 kJ/mol Reaksjonen går mot dannelse av fruktose-6-fosfat Fruktose-6-fosfat Glukose-6-fosfat 5 x 10-4 2 x 10-3
(298K)ln 0,41. = 2,2 kJ/mol. Reaksjonen går mot dannelse av glukose-6-fosfat. DG = DG’o + RT ln. = 2,2 kJ/mol + (8,3145 x 10-4 kJ/K/mol)(310K)ln. = 2,2 kJ/mol - 3,57 kJ/mol. = - 1,37 kJ/mol. Reaksjonen går mot dannelse av fruktose-6-fosfat. Fruktose-6-fosfat. Glukose-6-fosfat. 5 x x")
30
Bioenergetiske prinsipper II

31
Elektronforflytning utfører arbeid
Et batteri kan utføre arbeid når polene forbindes via f.eks en motor Denne åpner en vei slik at elektronene kan vandre mot den delen av batteriet som har størst elektronaffinitet. I metabolismen vandrer elektroner fra glukose til oksygen via en rekke elektrontransportører som da kan utføre arbeid e e e e e e Glukose O2

32
Elektrontransportkjeden
Elektroner vandrer gjennom elektrontransportkjeden mot O2 Elektronoverføringen er eksergon pga oksygens mere positive E’o Frigjort energi brukes til å skape en protongradient over membranen Protoner passerer tilbake gjennom membranen i en eksergon prosess som leverer energi til å fosforylere ADP til ATP

33
Redoksreaksjoner Reaksjoner hvor elektroner overføres fra et molekyl til et annet Tap av elektroner = oksidasjon Tilførsel av elektroner = reduksjon Elektronoverføringer er den egentlige basis for alt arbeid som utføres i en celle Organismer uten fotosyntese ekstraherer elektroner ut av næringsstoffer (mat) Fotosyntetiske organismer henter elektronene fra H2O ved hjelp av sollysets energi
 Fotosyntetiske organismer henter elektronene fra H2O ved hjelp av sollysets energi.")
34
Karbons mange oksidasjonstrinn

35
Karbons mange oksidasjonstrinn
Oksidasjoner involverer ikke nødvendigvis oksygen, det kan også være tap H (H+ + e-) Elektronoverføringer kan foregå på flere måter Direkte overføring: Fe 2+ + Cu 2+ <=> Fe 3+ + Cu+ Overføring av H: AH2 <=> A + 2e- + 2 H+ Overføring av hydridion: H- Oksygenering: R-CH3 + 1/2 O2 R-CH2-OH Reduserende ekvivalent = 2 e-
 Elektronoverføringer kan foregå på flere måter. Direkte overføring: Fe 2+ + Cu 2+ <=> Fe 3+ + Cu+ Overføring av H: AH2 <=> A + 2e- + 2 H+ Overføring av hydridion: H- Oksygenering: R-CH3 + 1/2 O2 R-CH2-OH. Reduserende ekvivalent = 2 e-")
36
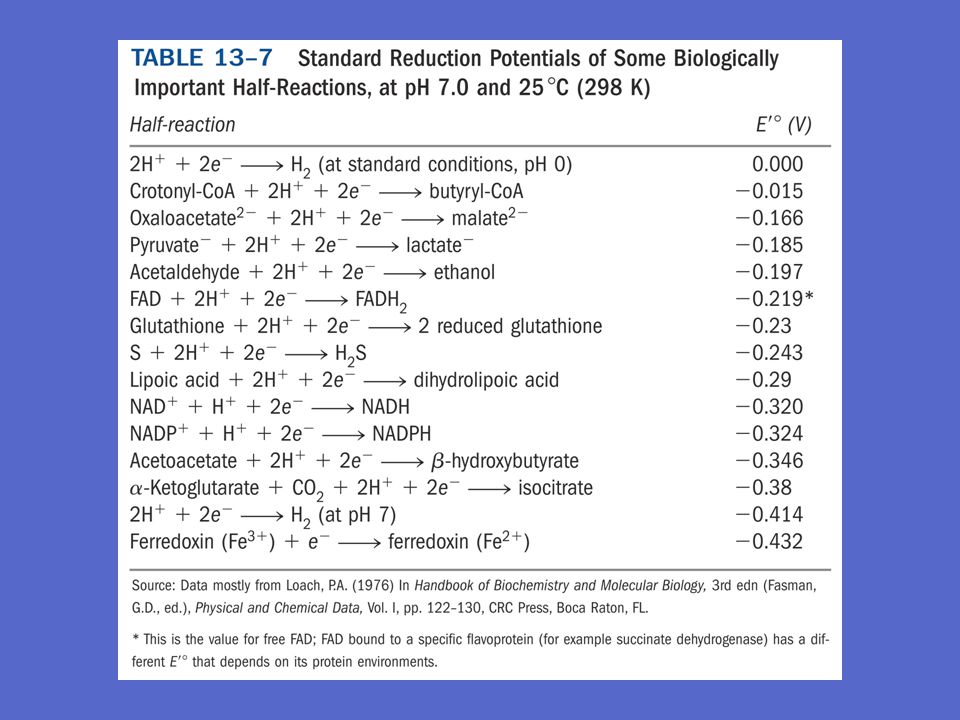
Standard reduksjonspotensialet Eo og fysiologisk SRP E’o
Standard reduksjonspotensialet, Eo angir en relativ affinitet for e- Biokjemien definerer standard reduksjonspotensialet ved fysiologiske forhold (298 K, pH 7) Elektroner vandrer til elektroden med det mest positiv E’o Referansen er: H+ + e /2 H2 , E’o = 0,00 V Både reduksjonspotensiale og konsentrasjon er med på å bestemme i hvilken retning elektronene vil vandre
 Elektroner vandrer til elektroden med det mest positiv E’o. Referansen er: H+ + e- 1/2 H2 , E’o = 0,00 V. Både reduksjonspotensiale og konsentrasjon er med på å bestemme i hvilken retning elektronene vil vandre.")
39
Reelt reduksjonspotensiale
Nernst likning: E = Eo + RT ln elektron mottaker nF elektron donor = Eo ln R = gasskonstanten T = o Kelvin n = antall elektroner overført/molekyle F = Faradays konstant elektron mottaker 0,026 V n elektron donor

40
Elektronoverføring representerer energi
Elektronoverføring utfører arbeid og representerer energi DE’o = E’o for elektron mottakeren - E’o for elektron donoren Eksempel: acetaldehyd + NADH + H etanol + NAD+ acetaldehyd + 2H+ + 2e etanol E’o = -0,197 V NAD+ + 2H+ + 2e NADH + H E’o = -0,320 V Elektronmottaker er reaksjonen med mest pos. E’o, dvs 1) DE’o = -0,197 - ( -0,320) = 0,123 V D’o er relatert til DG’o ved uttrykket DG’o = -nF DE’o DE’o = - 2(96,5 kJ/V x mol)(0,123 V) = - 23,7 kJ/mol Denne reaksjon vil altså gå mot etanol dannelse, når de deltakende forbindelsene er tilstede i 1M , men det har vi ikke i en levende celle
 DE’o = -0,197 - ( -0,320) = 0,123 V. D’o er relatert til DG’o ved uttrykket DG’o = -nF DE’o. DE’o = - 2(96,5 kJ/V x mol)(0,123 V) = - 23,7 kJ/mol. Denne reaksjon vil altså gå mot etanol dannelse, når de deltakende forbindelsene er tilstede i 1M , men det har vi ikke i en levende celle.")
41
Et mere realistisk eksempel
Hvis konsentrasjonen av acetaldehyd og NADH er 1M, mens konsentrasjonen av etanol og NAD+ er 0,1 M Eacetaldehyd = E’o + RT ln acetaldehyd nF etanol => -0,167 V ENADH = E’o + RT ln NAD+ nF NADH => -0,350 V DE = -0,167 V - (- 0,350 V) = 0,183 V DG = -nF DE = -2( 96,5 kJ/Vmol)(0,183V) = - 35,3 kJ/mol Viser kraftig forskyvning i retning av produktdannelse
 = 0,183 V. DG = -nF DE. = -2( 96,5 kJ/Vmol)(0,183V) = - 35,3 kJ/mol. Viser kraftig forskyvning i retning av produktdannelse.")
42
Elektronoverføring i metabolismen skjer i mange små trinn
Fullstendig oksidasjon av glukose har en DG’o = kJ/mol Oksidasjonen skjer i mange trinn av passe størrelse for å ta vare på mest mulig energi, typisk den energimengde som kreves for å danne én ATP Elektronoverføringen skjer bare unntaksvis direkte til ATP Mange elektronbærende kofaktorer deltar i nedbrytningen av glukose

43
Elektronbærende kofaktorer
NAD+, NADP+, FMN, FAD er vannløselige kofaktorer NAD+, NADP+ assosieres relativt løst til sine enzymer og kan vandre mellom proteiner FMN, FAD assosieres tett til sine enzymer, flavoproteinene og betraktes som prostetiske grupper. Forlater dem ikke Quinoner (ubiquinon, plastoquinon) er lipidløselige kofaktorer og fungerer i membraner Jern-svovl proteiner har tett bundne kofaktorer, prostetiske grupper Cytokromene har tett bundne kofaktorer, prostetiske grupper
 er lipidløselige kofaktorer og fungerer i membraner. Jern-svovl proteiner har tett bundne kofaktorer, prostetiske grupper. Cytokromene har tett bundne kofaktorer, prostetiske grupper.")
44
NAD+ og NADP+

45
NAD+ og NADP+ NAD+ og NADH er kofaktorer for enzymene i katabolismen
Vanlig konsentrasjon totalt for disse er 10 mM Det er mest NAD+ noe som favoriserer overføring av H- fra substrater NADP+ og NADPH er kofaktorer for enzymene i anabolismen Vanlig konsentrasjon totalt for disse er 1 mM Det er mest NADPH noe som favoriserer syntese av nye molekyler De fungerer alle som kofaktorer for enzymer av typen oksidoreduktaser eller dehydrogenaser

46
Spesifisitet i valg av kofaktor

47
Spesifisitet i valg av kofaktorer

48
FAD og FMN

49
FAD og FMN Disse kan ta imot én eller to elektroner i form av hydrogen atomer FAD + H = FADH + H = FADH2 Disse flavinnukleotider er svært tett bundet til sine enzymer og anses som prostetiske grupper På grunn av sin tette assosiering har disse varierende reduksjons-potensiale

Liknende presentasjoner

















>")
>")

