Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
Superhelikstopologi: Supercoiling av DNA
Lk = Tw + Wr Lk: linking number, det topologiske tvinningstall. Antall ganger en DNA-tråd tvinner seg rundt den andre. Konstant for et kovalent lukket molekyl eller et molekyl hvor endene er immobilisert Tw: twist, antall fullstendige omdreininger en tråd gjør rundt heliksaksen. Positiv for høyredreiende heliks. B-DNA: 10.4 Wr: writhing number, antall omdreininger dupleksaksen gjør rundt superheliksaksen. ”Supercoiltvinningstallet”

2
Supercoiled DNA: fjerning av supercoils ved separasjon av DNA-trådene
Relationship of linking number, twist and writhe of closed circular forms of DNA. Closed DNA circles can be made by formation of covalent 5'–3' phosphodiester bonds on each strand of a linear molecule. For a linear molecule with 36 helical turns, the linking number of this unconstrained state (Lk0) is 36. Closure into an unconstrained planar circle, as shown on the left side of the figure, produces a molecule with twist (Tw) = 36 and writhe (Wr) = 0. If the number of helical turns is altered before closure, the DNA molecule adopts a supercoiled conformation. On the right side of the figure, four helical turns are removed from the molecule, reducing the linking number (Lk) to 32. For simplicity, the figure shows all unwinding partitioned as Wr, although such changes are usually partitioned between Tw and Wr. Unwinding of helical turns produces negatively supercoiled DNA (or -DLk) as shown, whereas the inclusion of additional turns produces positively supercoiled DNA. For DNA with -DLk in the interwound form, the superhelical turns are right-handed. Note that separation of DNA strands removes negative supercoils (equivalent to the addition of positive supercoils). Encyclopedia of Life Sciences / Nature Publishing Group ©2001 Macmillan Publishers Ltd. Reg.No England (except where otherwise stated)
 is 36. Closure into an unconstrained planar circle, as shown on the left side of the figure, produces a molecule with twist (Tw) = 36 and writhe (Wr) = 0. If the number of helical turns is altered before closure, the DNA molecule adopts a supercoiled conformation. On the right side of the figure, four helical turns are removed from the molecule, reducing the linking number (Lk) to 32. For simplicity, the figure shows all unwinding partitioned as Wr, although such changes are usually partitioned between Tw and Wr. Unwinding of helical turns produces negatively supercoiled DNA (or -DLk) as shown, whereas the inclusion of additional turns produces positively supercoiled DNA. For DNA with -DLk in the interwound form, the superhelical turns are right-handed. Note that separation of DNA strands removes negative supercoils (equivalent to the addition of positive supercoils). Encyclopedia of Life Sciences / Nature Publishing Group ©2001 Macmillan Publishers Ltd. Reg.No England (except where otherwise stated)")
3
Sirkulære DNA-molekyler med forskjellig grad av supercoil

4
To topologisk ekvivalente supercoilformer og en biologisk variant

5
Interkalerende stoffer kontrollerer supercoiling ved å opptvinne DNA
Etidiumbromid Akridinoransje Proflavin

6
Variasjon i supercoil i en negativt coiled superheliks med økende etidiumkonsentrasjon

7
Elektroforetisk analyse av supercoil i DNA
Skjematisk elektroforese av en DNA-prøve med og uten interkalator. Prøven inneholder nicked DNA (N), lineært DNA (L) og negativt supercoiled DNA (SC) Elektroforese av DNA med forskjellig grad av supercoil ved forskjellige konsentrasjoner av interkalator 2D-elektroforese av topoisomerer med fra svært negativ til svakt positiv supercoiling. D1: uten interkalator, D2: 20 mg/ml klorokin Measurement of linking number by gel electrophoresis. (a) Schematic illustration of a DNA sample separated by electrophoresis through an agarose gel with and without an intercalator. DNA isolated from bacterial cells contains molecules with different topology: some have their backbones unbroken and are negatively supercoiled (SC), some have one strand broken or ‘nicked’ (N) and some have both strands broken to produce a linear molecule (L). Note that the supercoiled DNA consists of a Gaussian distribution of different topoisomers. Upon addition of intercalator, the migration of intact molecules is altered, but that of nicked and linear molecules is not changed. (b) Relaxation of plasmid DNA in the presence of varying concentrations of intercalator produces samples containing topoisomers at different levels of supercoiling. Utilization of multiple gels with different concentrations of intercalator allows measurement of DLk. For each sample, average superhelical density (s) is shown above the lane. Note that in each gel, samples can have positively or negatively supercoiled topoisomers. The inclusion of intercalator in the running buffer alters the electrophoretic mobility of all topoisomers equivalently. Superhelical density can be measured for experimental samples (‘native’) by comparison with those of known s. (c) Two-dimensional agarose gel electrophoresis of topoisomers ranging from high negative s to moderate positive s. A DNA sample is loaded in a single well in a large agarose gel and electrophoresis is performed under specific conditions (usually without intercalator) in direction D1. After soaking of the gel in buffer containing intercalator, electrophoresis is continued in direction D2 (90° to D1). The gel shown contained 20 mg mL-1 chloroquine during the second electrophoresis, resulting in all topoisomers having positive s. Deviation of topoisomers from a smooth curve indicates that structural transitions reduced their negative s during the first direction of electrophoresis. Spots marked ‘N’ and ‘L’ indicate the position of migration of ‘nicked’ and ‘linear’ DNA molecules, respectively
, lineært DNA (L) og negativt supercoiled DNA (SC) Elektroforese av DNA med forskjellig grad av supercoil ved forskjellige konsentrasjoner av interkalator. 2D-elektroforese av topoisomerer med fra svært negativ til svakt positiv supercoiling. D1: uten interkalator, D2: 20 mg/ml klorokin. Measurement of linking number by gel electrophoresis. (a) Schematic illustration of a DNA sample separated by electrophoresis through an agarose gel with and without an intercalator. DNA isolated from bacterial cells contains molecules with different topology: some have their backbones unbroken and are negatively supercoiled (SC), some have one strand broken or ‘nicked’ (N) and some have both strands broken to produce a linear molecule (L). Note that the supercoiled DNA consists of a Gaussian distribution of different topoisomers. Upon addition of intercalator, the migration of intact molecules is altered, but that of nicked and linear molecules is not changed. (b) Relaxation of plasmid DNA in the presence of varying concentrations of intercalator produces samples containing topoisomers at different levels of supercoiling. Utilization of multiple gels with different concentrations of intercalator allows measurement of DLk. For each sample, average superhelical density (s) is shown above the lane. Note that in each gel, samples can have positively or negatively supercoiled topoisomers. The inclusion of intercalator in the running buffer alters the electrophoretic mobility of all topoisomers equivalently. Superhelical density can be measured for experimental samples (‘native’) by comparison with those of known s. (c) Two-dimensional agarose gel electrophoresis of topoisomers ranging from high negative s to moderate positive s. A DNA sample is loaded in a single well in a large agarose gel and electrophoresis is performed under specific conditions (usually without intercalator) in direction D1. After soaking of the gel in buffer containing intercalator, electrophoresis is continued in direction D2 (90° to D1). The gel shown contained 20 mg mL-1 chloroquine during the second electrophoresis, resulting in all topoisomers having positive s. Deviation of topoisomers from a smooth curve indicates that structural transitions reduced their negative s during the first direction of electrophoresis. Spots marked ‘N’ and ‘L’ indicate the position of migration of ‘nicked’ and ‘linear’ DNA molecules, respectively.")
8
Sekvensindusert bøying av DNA

9
Proteinindusert DNA-bøying
Role of architectural transcription factors. (a) The region upstream of a hypothetical gene, with the start of the transcribed region shown as a solid bar. A, B and C represent sites for binding of regulatory proteins. In this example, B represents a binding site for an architectural transcription factor. (b) Binding of factor B (shaded oval), resulting in bending of the chromatin. As a result, binding sites for A and C are brought into close proximity. (c) Binding of factors A and C then allows a complex of transcription factors to be built up (ovals and circles), such that transcription of the hypothetical gene is initiated (arrow).
 The region upstream of a hypothetical gene, with the start of the transcribed region shown as a solid bar. A, B and C represent sites for binding of regulatory proteins. In this example, B represents a binding site for an architectural transcription factor. (b) Binding of factor B (shaded oval), resulting in bending of the chromatin. As a result, binding sites for A and C are brought into close proximity. (c) Binding of factors A and C then allows a complex of transcription factors to be built up (ovals and circles), such that transcription of the hypothetical gene is initiated (arrow).")
10
Elektroforetisk analyse av bøyd DNA

11
Topoisomeraser Type I topoisomeraser virker ved å innføre forbigående enkelttrådsbrudd i DNA Type II topoisomeraser virker ved å innføre forbigående dobbelttrådsbrudd i DNA

12
Virkning av topoisomerase I på supercoiled DNA
Negativt supercoiled, nativt SV40-DNA Behandlet med topo I i 5 min Behandlet med topo I i 30 min

13
Topo I: en pekepinne mot mekanismen

14
Kovalent enzym-DNA-kompleks som intermediat

15
Topo IA: et forslag til mekanisme

16
Topo IB: et forslag til mekanisme

17
Topo II kutter begge tråder i en dupleks, lar dupleksen passere gjennom bruddstedet og forsegler bruddet

18
Topo II: Et forslag til mekanisme

19
Topo IA, Topo IB og Topo II: Mekanismer
Figure 4 Mechanistic models of topoisomerases. (a) Type IA topoisomerases. (b) Type IB topoisomerases (based on Stewart et al. (1998). (c) Type II topoisomerases. Separate domains or monomers are shown in red and blue, active-site tyrosines in yellow, and ATP as yellow ellipses. See text for mechanistic details.
 Type IA topoisomerases. (b) Type IB topoisomerases (based on Stewart et al. (1998). (c) Type II topoisomerases. Separate domains or monomers are shown in red and blue, active-site tyrosines in yellow, and ATP as yellow ellipses. See text for mechanistic details.")
20
Utvalgte topoisomerasers egenskaper

21
Topo II-inhibitorer

22
Restriksjonsendonukleaser
Type I: Endonuklease og metylase i samme polypeptid. Spalting på tilfeldig(?) sete minst 1000 bp fra gjenkjenningssetet Type II: Endonuklease og metylase separate enzymer. Spalter spesifikt i gjenkjenningssekvensen Type III: Endonuklease og metylase i samme polypeptid. Spalter bp fra gjenkjenningssekvensen
 sete minst 1000 bp fra gjenkjenningssetet. Type II: Endonuklease og metylase separate enzymer. Spalter spesifikt i gjenkjenningssekvensen. Type III: Endonuklease og metylase i samme polypeptid. Spalter bp fra gjenkjenningssekvensen.")
23
Palindrome gjenkjenningsseter

24
DNA polymeraser: med et templat, en primer og dNTP produseres en komplementær tråd

25
Dideoksynukleotider (ddNTP) vil terminere voksende kjeder
 vil terminere voksende kjeder")
26
Terminering med ddNTP styres av templatsekvensen

27
Separasjon av de forskjellige fragmentene i reaksjonsblandingen ved elektroforese

28
Figure 7-15 Autoradiograph of a sequencing gel.
Page 179

29
Sequencing with labelled terminators
GATC ddATP ddCTP ddGTP ddTTP

30
Litt sekvens fra et automatisert DNA-sekvenseringsinstrument
Og en animering av DNA-sekvensering

31
Polymerase families Family A Family B Family C Family X
DNA polymerase I DNA polymerase Family B DNA polymerase II DNA polymerases , , Archae polymerases (Vent, Pfu, Pwo…) Family C DNA polymerase III Family X DNA polymerase Terminal transferase
 Family C. DNA polymerase III. Family X. DNA polymerase Terminal transferase.")
32
Family A DNA polymerases
C 5’-3’ exo domain 3’-5’ exo domain Polymerase domain Polymerase 5’-3’ 3’-5’ Pol E. coli polymerase I + Klenow fragment - T7 DNA polymerase Taq polymerase

33
Hvorfor varmestabile DNA polymeraser for sekvensering?
Sekvensreaksjonen kan kjøres flere ganger på samme templat, dette øker sensitiviteten slik at mindre templat kan brukes Ved høye temperaturer er det mindre problemer med renaturering og dannelse av sekundærstrukturer i DNA

34
Figure 7-17 Genome sequencing strategies.
Page 180

35
DNA polymeraser: innføring av et nytt nukleotid

36
Korrekturlesning: DNA polymerase I fjerner et feilinkorporert nukleotid

37
5’-3’-eksonukleasedomenet i DNA polymerase I fjerner nukleotider i 5’ –enden av et nick

38
Klenow-fragmentet av DNA polymerase I i kompleks med DNA

39
Klenow-fragmentets to virkemåter

40
Nick-translasjon katalysert av DNA polymerase I

41
DNA polymeraser fra E.coli

42
Polymerase families Family A Family C Family X Family B
DNA polymerase I DNA polymerase Family B DNA polymerase II DNA polymerases , , Archae polymerases (Vent, Pfu, Pwo…) Family C DNA polymerase III Family X DNA polymerase Terminal transferase
 Family C. DNA polymerase III. Family X. DNA polymerase Terminal transferase.")
43
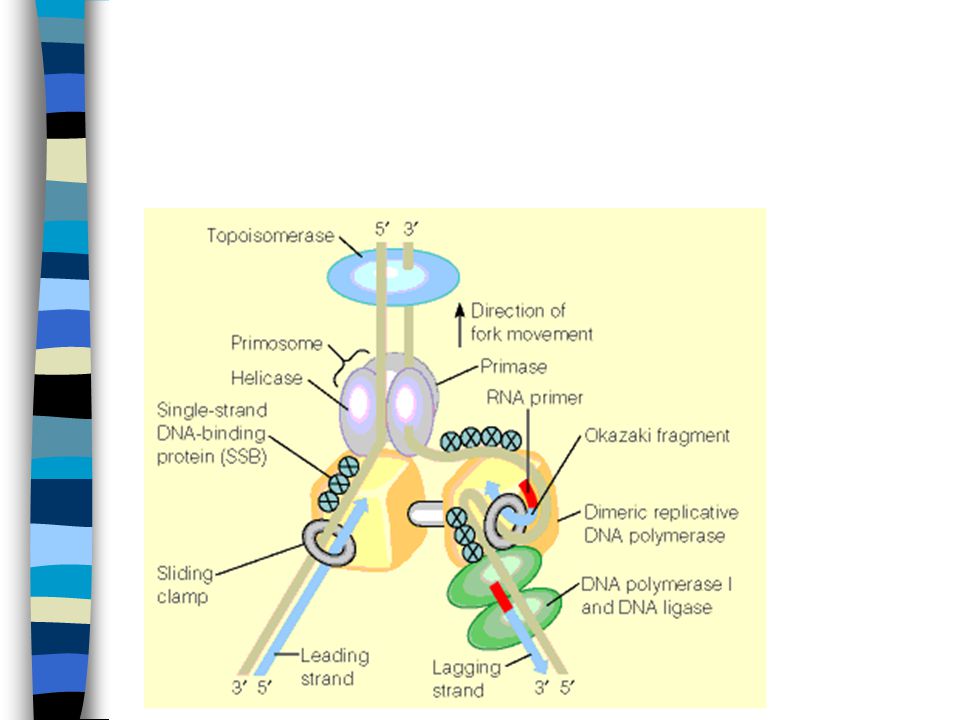
DNA polymerase III: En komplisert maskin

44
DNA polymerase III: En komplisert maskin

45
”Clamp loader”-komplekset monterer β-subenheten på DNA

46
β-subenheten i mer detalj
PolI-mutanter kan replikere DNA – pol I er ikke replikasen. Men mutanten er UV-følsom – pol I deltar i reparasjon. Fjerner også RNA-primere og fyller inn

Liknende presentasjoner









 happens in the database: : data.>")









