Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
Nukleinsyrenes struktur
Primær- og sekundærstruktur

2
Nukleinsyrestruktur Nukleinsyrestruktur beskrives på tre nivåer:
Primærstruktur forteller om nukleotidsekvensen Sekundærstruktur forteller hvordan polynukleotidet er kveilet opp Tertiærstruktur forteller hvordan polynukleotidet er assosiert med proteiner og hvordan det foreligger i kromatinet Her skal vi bare beskjeftige oss med det andre nivået

3
DNA er en dobbel heliks DNA består av to poly-nukleotidtråder som danner en dobbelheliks Heliksen er høyredreid Det er ca. 10 nukleotider pr heliksomdreining Hver omdreining er 36 Å

4
DNA er en dobbel heliks Polynukleotidtrådene holdes sammen av hydrogenbindinger mellom basene A T og C G Basene sitter i heliksens indre => minst mulig kontakt mellom de hydrofobe basene og det hydrofile miljøet rundt Deoksyribosefosfatryggraden danner heliksens overflate => god kontakt mellom de negativt ladete og polare gruppene

5
Stabilisering av dobbel heliksen
To typer krefter stabiliserer heliksen Hydrogenbindinger mellom polynukleotid-trådene holder de to trådene sammen = tverrgående krefter Hydrofobe interaksjoner mellom basene i heliksens lengderetning = langsgående krefter Basestacking er viktigst for stabilisering

6
Heliksstruktur Major groove og minor groove
Baseparene ligger i samme plan Hvert basepar forlenger heliksen med 3.4 Å Hver omdreining forlenger heliksen med 36 Å og er på 10,5 basepar i vandig miljø Figur a og c

7
5’ 3’ Heliksen DNA trådene har motsatt orientering, de er antiparallelle 5’ ’ 3’ ’ DNA trådene er komplementære Baseparringen => at de to trådene ikke er identiske men hver av dem gir full informasjon om nukleotidsekvensen i den andre 3’ 5’

8
Semikonservativ replikasjon
Ved celledeling syntetiseres fullt kopisett av DNA DNA trådene skilles og fungerer som templat for syntese av komplementærtråden Hver dattercelle får en ”gammel” DNA tråd og en ”ny” Systemet med dobbeltrådet DNA er veldig sikkert, dvs få feil forblir uoppdaget Skade på en tråd repareres vha informasjonen på den andre tråden Feil = mutasjoner, noen få => evolusjon Alle organismer unntatt noen virus benytter dette systemet Figur 10-17

9
Tre heliksstrukturer A B Z B-strukturen er den vanlige i vandig miljø
A-strukturen er mere kompakt, finnes i ikke-vandig miljø og neppe in vivo Z-strukturen er venstredreid og mere langstrakt. Finnes in vivo men funksjon er ukjent Major groove er nesten borte Minor groove er trang og dyp Spesielt ved GC-rike sekvenser Figur 10-19

10
A-, B- og Z-helikser

11
Spesielle strukturer er involvert i genregulering
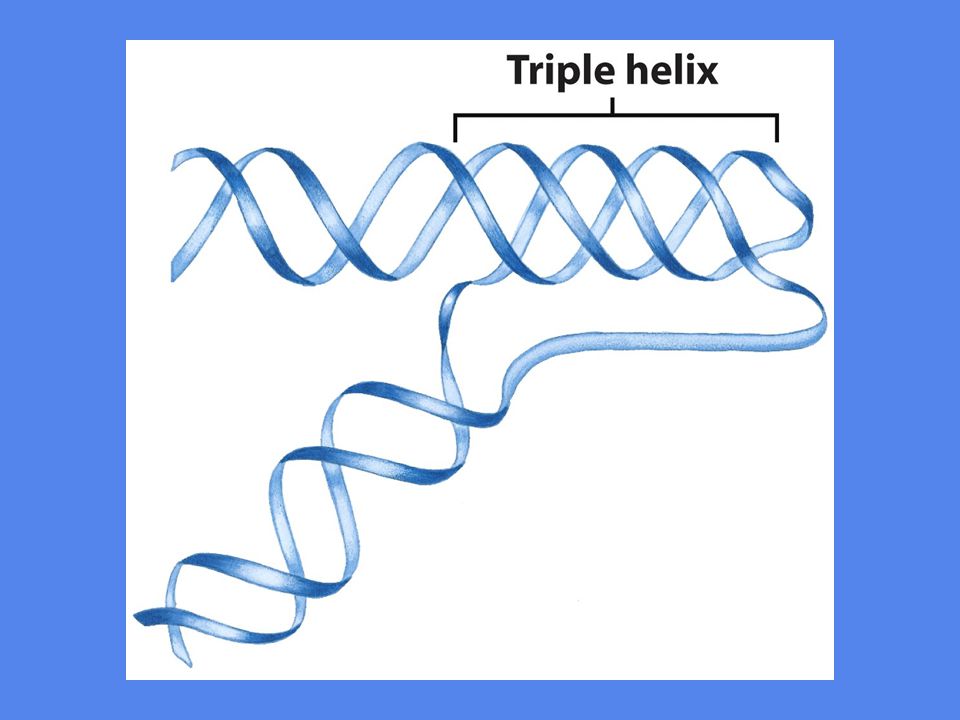
Palindrom: gjentakelse av sekvensen i motsatt retning på motsatt tråd Er internt komplementære Speilkopi: gjentakelse av sekvensen i motsatt retning på samme tråd 3-4 adeniner => bøy på DNA Trippelheliksen

12
Palindromer F.eks. rotor
Palindrom = ord som er likt når det leses forfra og bakfra F.eks. rotor

13
Hårnålstruktur

15
Trippelheliksen H-DNA struktur
Forekommer i områder hvor sekvensen er alternerende T og C eller A og G I denne strukturen er to av trådene dannet av bare pyrimidiner, C og T, mens den tredje tråden består av bare puriner, G og A Ofte i områder involvert i regulering av DNA-aktivitet: replikasjon, transkripsjon, rekombinasjon

17
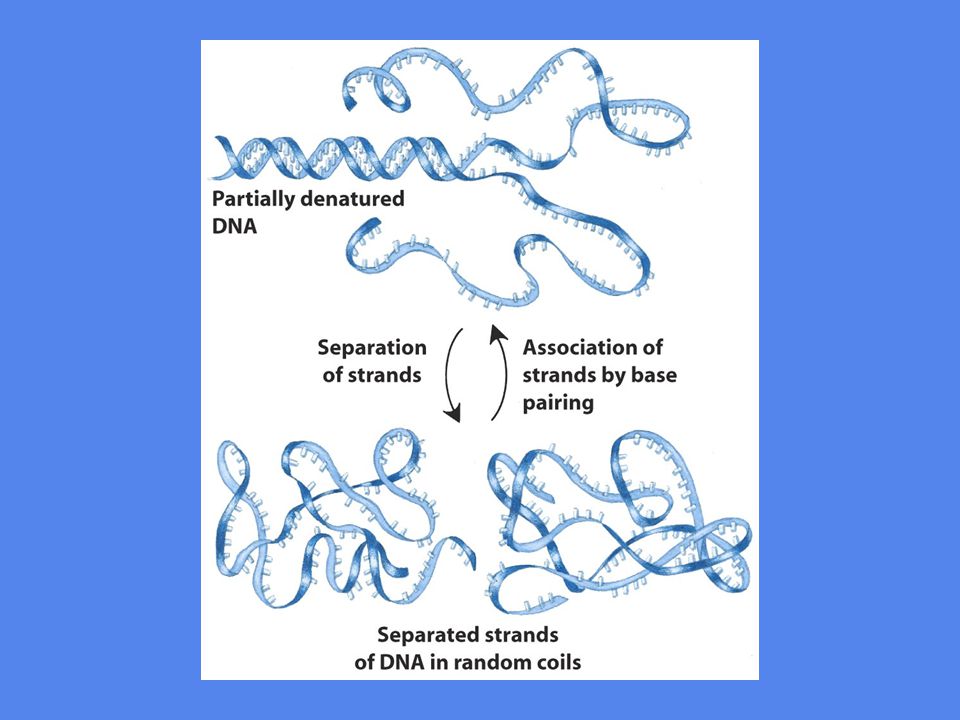
Denaturering DNA trådene holdes sammen av ikke-kovalente bindinger
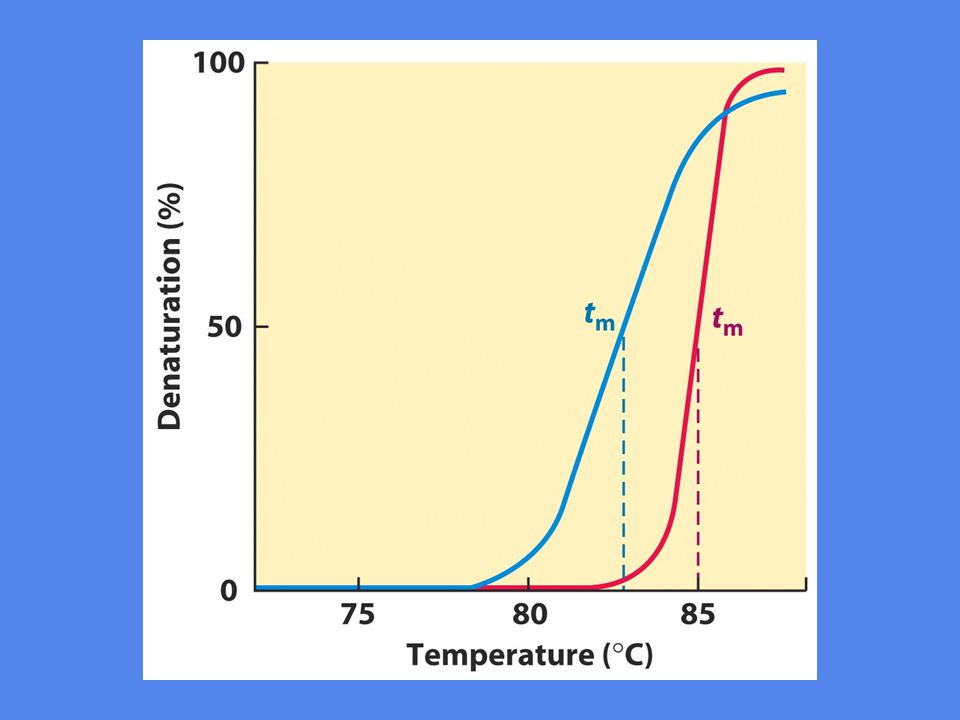
Ekstrem pH eller høy temperatur kan bryte disse bindingene Ved denaturering løses strukturer opp og DNA trådene foreligger som ”random coils” Denaturering medfører økt UV-absorpsjon Figur 10-29

18
Hyperkrom effekt Absorpsjon av lys i UV-området er størst for frie nukleotider. Enkelttrådet DNA/RNA har lavere absorpsjon på grunn av ”base stacking” Dobbelttrådet DNA/RNA har lavest absorpsjon fordi både ”base stacking” og baseparing reduserer den.

19
Varmedenaturering Denatureringstemperaturen avhenger av flere forhold:
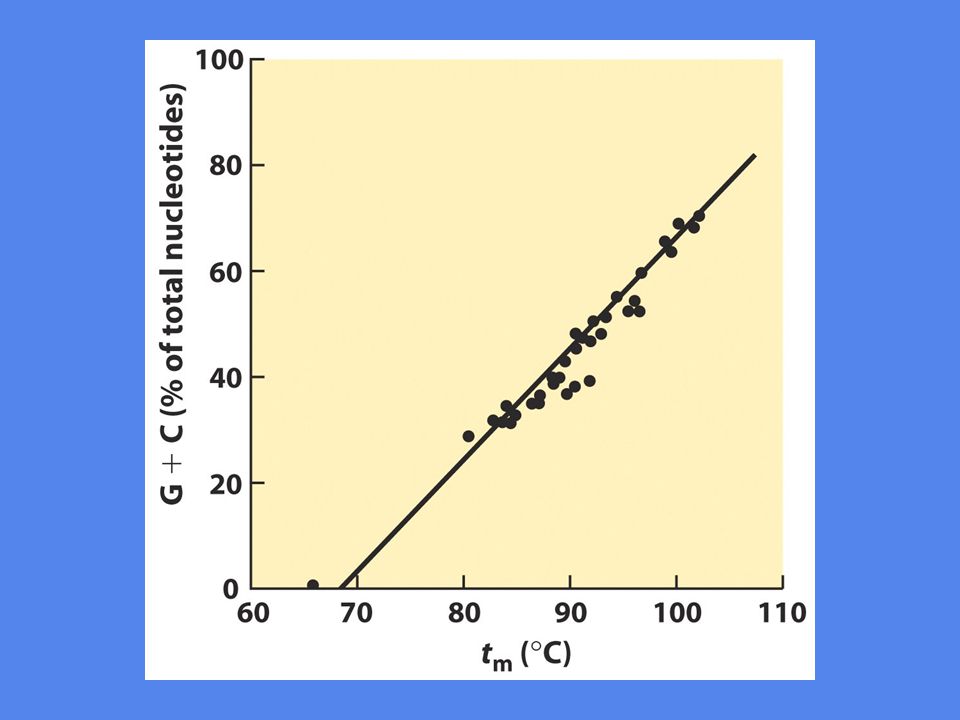
Viktigst er nukleotidsekvensen Det krever mere energi å bryte et CG basepar enn et AT basepar, fordi CG har tre hydrogenbindinger mens AT bare har to Lengden på DNA molekylet spiller også inn, mest relevant in vitro In vivo foregår denaturering i forbindelse med replikasjon og transkripsjon Denatureringstemperatur er høyest for RNA:RNA, lavere for RNA:DNA og lavest for DNA:DNA. Årsaken er ikke kjent.

22
Renaturering Denaturering kan være hel eller delvis, den motsatte prosessen heter renaturering Renaturering kalles også ”annealing” Renaturering er en spontan prosess som foregår når temperaturen eller pH-forholdene tillater det Renaturering er en rask prosess hvis de to DNA trådene har bare et lite område intakt heliks - ellers kan det mere tid fordi trådene må finne riktig start Dobbeltrådete strukturer kan dannes mellom to DNA tråder, innen eller mellom to RNA tråder eller mellom en DNA og en RNA tråd

25
RNA Det er tre forskjellige typer RNA Messenger RNA = mRNA
Transfer RNA = tRNA Ribosomalt RNA = rRNA

26
Messenger RNA I en prosess som kalles transkripsjon kopieres DNA til messenger RNA Både celler med og uten kjerne, eukaryote og prokaryote, bruker mRNA som bindeledd mellom DNA og proteinsyntesemaskineriet Messenger RNA er templat ved proteinsyntese i ribosomene I eukaryote celler er mRNA monosistronisk, dvs det bærer koden for et protein Dette mRNA har en lengde som minimum omfatter sekvensen til det proteinet det koder for, dvs den nukleotidsekvens som kalles for åpen leseramme (open reading frame = ORF) I den åpne leserammen er det tre nukleotider for hver aminosyre som skal defineres ORF
 I den åpne leserammen er det tre nukleotider for hver aminosyre som skal defineres. ORF.")
27
Messenger RNA Det betyr at for et protein med 100 aminosyrer trengs et mRNA med minst 300 nukleotider mvRNA er mye lengre i virkeligheten På hver side av den åpne leserammen finnes områder som har med regulering å gjøre, f eks kontroll av proteinsyntesehastighet ORF

28
Messenger RNA I prokaryote celler bærer mRNA ofte flere åpne leserammer Dette kalles polysistronisk mRNA ORF ORF ORF 3

29
RNA struktur Både rRNA, tRNA og mRNA produseres ved transkripsjon
Alle RNA er enkelttrådete RNA danner forskjellige strukturer ved å baseparre med komplementære områder RNA kan danne høyredreide heliksstrukturer av A- og Z-typen

30
Andre nukleotidfunksjoner
Utover å være byggesteiner i DNA og RNA spiller nukleotidene en viktig rolle som: 1) Energivaluta i cellen 2) Delkomponent i mange koenzymer 3) Signalmolekyler
 Energivaluta i cellen. 2) Delkomponent i mange koenzymer. 3) Signalmolekyler.")
32
ATP er cellens energivaluta

33
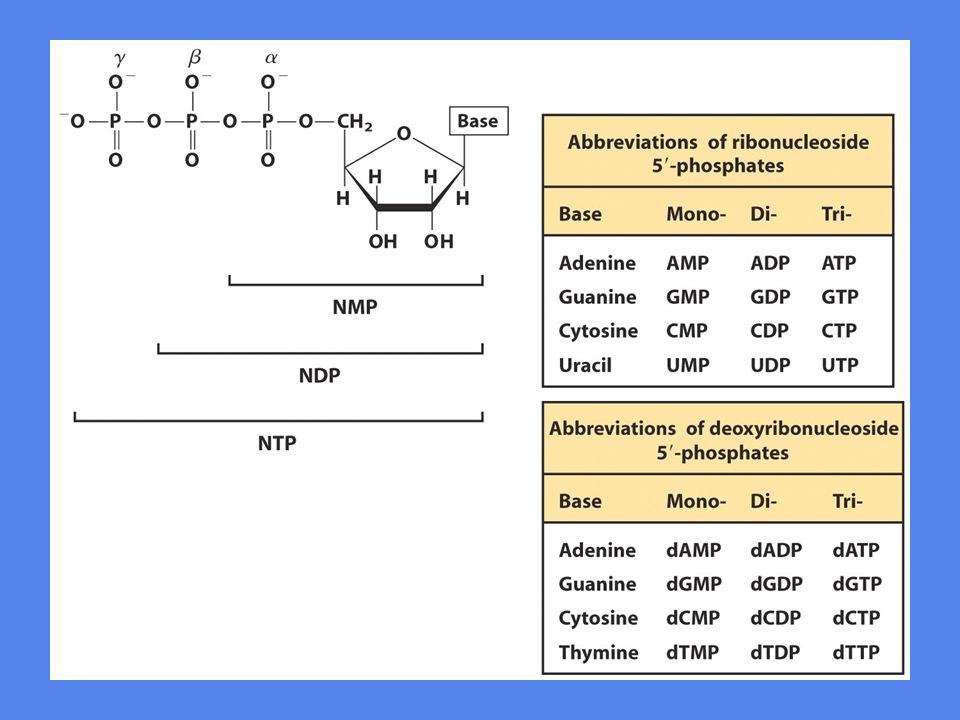
Cellens energivaluta Trinukleotidet ATP er viktigste energibærer
GTP, UTP og CTP deltar i spesielle reaksjoner Energien finner vi anhydrid-bindingene mellom fosfatgruppene som sitter på C-atom 5’ i ribosen Fosfatgruppene betegnes a,b,g regnet fra C-5’ Esterbindingen frigjør ca 14 kJ/mol ved hydrolyse Anhydridbindingene frigjør ca 30 kJ/mol ved hydrolyse

34
Nukleotidtrifosfater som energibærere
Hvorfor bruker cellen så kompliserte molekyler som energibærere? Binding av ATP til et enzym fører til dannelse av mange ikke-kovalente bindinger og dermed frigjøring av mye bindingsenergi Bindingsenergien kommer i tillegg til hydrolyseringsenergien og gjør dermed ATP til en mere effektiv energidonor

35
Fosfonukleotider som komponenter i koenzymer

36
Fosfonukleotider som komponenter i koenzymer

37
Fosfonukleotider som komponenter i koenzymer

38
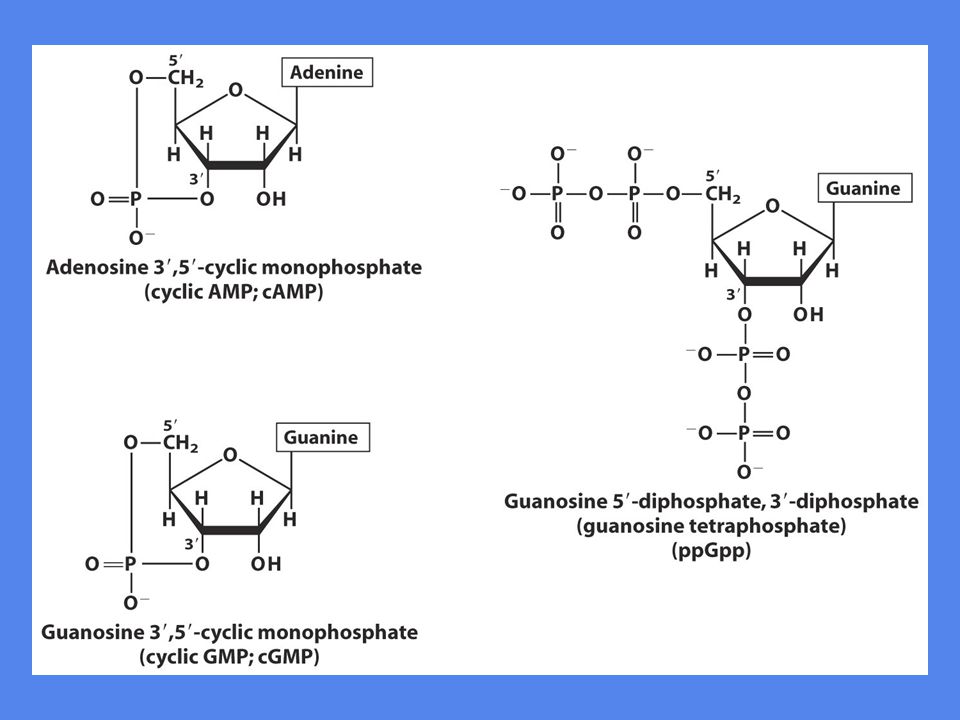
Fosfonukleotider som signalmolekyler
To viktige intracellulære signalmolekyler: cAMP og cGMP Fungerer som bærere av signaler generert ved cellemembranen cAMP er viktig i alle celler cAMP dannes av enzymet adenylyl cyclase som er assosiert med innsiden av cellemembranen Bakterier har signalmolekylet guanosintetrafosfat ppGpp

Liknende presentasjoner



















 og usynlig (radiofrekvens) energi.>")

