Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
FORELESNINGSPLAN DNA replikasjon Prokaryot: 2t, 16/2 Eukaryot: 2t, 25/2 DNA reparasjon t, 1/3 Erik Boye Avdeling for Cellebiologi Det norske Radiumhospital HF

2
Det sentrale problemet med cellesyklus
VEKST CELLEDELING Hver dattercelle må få tildelt alle essensielle komponenter, som mitokondrier, ribosomer, organeller, etc. Disse er til stede i flere kopier, og da er det tilstrekkelig med en tilfeldig fordeling mellom dattercellene Hver dattercelle må få et fullstendig sett av alt kromosomalt DNA. Kromosomene må derfor bli duplisert, og de to kopiene må segregeres til hver datter. Denne prosessen må være helt nøyaktig og presist regulert.

3
How does DNA replicate? Watson & Crick (1953) Nature 171: 737-738.
“It has not escaped our notice that the specific pairing we have postulated immediately suggests a possible copying mechanism for the genetic material.” Parent DNA New DNA KEY Parent double helix Possible modes of DNA replication (a) dispersive (b) semi-conservative (c) conservative
 dispersive. (b) semi-conservative. (c) conservative.")
4
The Meselson-Stahl experiment (1958) Meselson and Stahl (1958) Proc
The Meselson-Stahl experiment (1958) Meselson and Stahl (1958) Proc. Natl. Acad. Sci. USA 44: E. coli cells were grown for many generations in ‘heavy’ medium (containing 15N, a heavy isotope of nitrogen). Cells were then transferred to ‘light’ medium (containing 14N). DNA samples were analysed by density gradient centrifugation. Heavy medium Light medium, 1 generation Light medium, 2 generations Inter- pretation:
 Meselson and Stahl (1958) Proc. Natl. Acad. Sci. USA 44: E. coli cells were grown for many generations in ‘heavy’ medium (containing 15N, a heavy isotope of nitrogen). Cells were then transferred to ‘light’ medium (containing 14N). DNA samples were analysed by density gradient centrifugation. Heavy medium. Light medium, 1 generation. Light medium, 2 generations. Inter- pretation:")
5
Hvilke oppgaver må løses under DNA replikasjon?
Kopiering av alle DNA-sekvenser Kopiering kun EN gang pr celledeling, hverken mer eller mindre. Nøyaktig kopiering gir genetisk stabilitet. Hvordan oppnå dette? Definerte startpunkter (origins) Finregulering av initiering Nøyaktige polymeraser Reparasjonsmekanismer Presis segregering
 Finregulering av initiering. Nøyaktige polymeraser. Reparasjonsmekanismer. Presis segregering.")
6
Mechanism of DNA synthesis
OH 5’ 3’ triphosphate 5’ deoxyribose phosphate Bases adenine thymine cytosine guanine hydrogen bond release of pyrophosphate OH 3’ hydrolysis 5’ 3’

7
Replikasjonsgaffel ?

8
DNA polymeraser kan bare syntetisere 5’-3’.
Hvordan går det med to antiparallelle tråder?

9
Hvordan replikere en DNA-dupleks med enzymer som bare virker i én retning?
Vist eksperimentelt av Reiji Okazaki i 1968

10
Syntese av Okazaki-fragment og ny priming

11
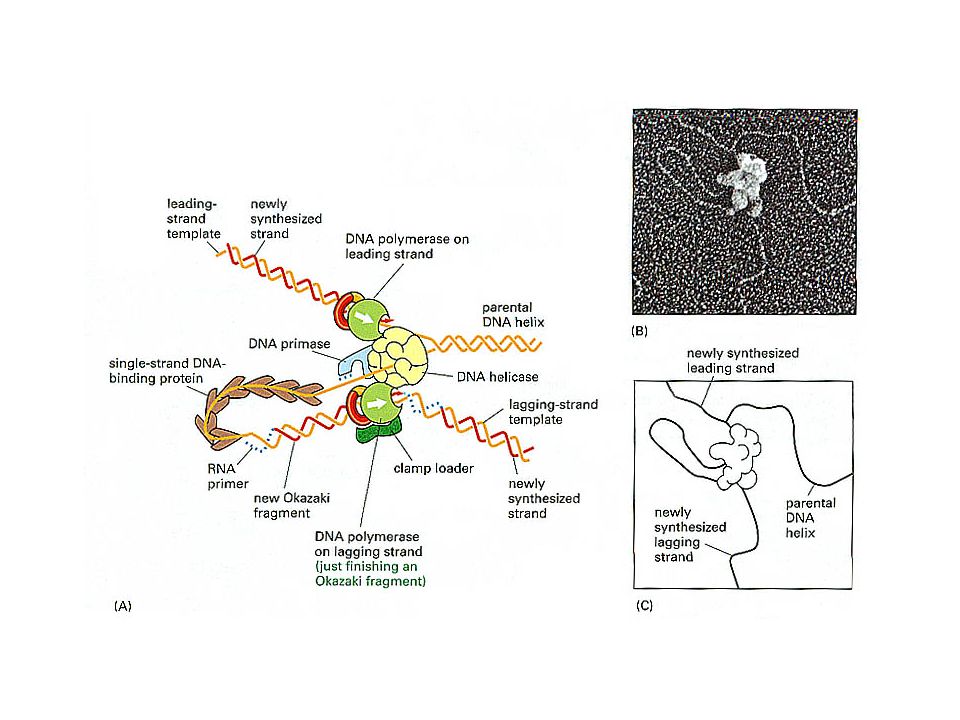
Semidiskontinuerlig replikasjon
Elektronmikroskopisk bilde av replikerende D. melanogaster-DNA som bekrefter Okazaki-modellen (Kreigstein & Hogness 1974)
")
12
Sammensetning av primosomet

13
DNA polymerase III: En komplisert maskin

14
DNA polymerase III: En komplisert maskin

15
Andre proteiner som deltar i replikasjonen
DNA ligase - Covalently closes nicks in double-stranded DNA. Primase - the enzyme responsible for catalyzing synthesis of RNA primers. Clamps and clamp loaders - Protein from the DNA polymerase III holoenzyme complex holds the polymerase to the DNA. A multisubunit entity called the complex functions as the "clamp loader". Single-strand DNA-binding (SSB) proteins – stabilizes ssDNA in the replication fork Helicases – unwinds DNA by catalyzing the ATP-dependent unwinding of double-strand DNA. E. coli contains at least 6 different helicases--some involved in DNA repair and others in conjugation. The principal helicase in DNA replication is DnaB, which interacts with DnaG and other proteins to form the primosome. Topoisomerases - Bidirectional replication of the circular E. coli chromosome unwinds about 100,000 base pairs per minute. Relief of this torsional stress is essential for DNA replication to occur. Topoisomerases are enzymes with a "swivel" mechanism that can relieve this stress.
 proteins – stabilizes ssDNA in the replication fork. Helicases – unwinds DNA by catalyzing the ATP-dependent unwinding of double-strand DNA. E. coli contains at least 6 different helicases--some involved in DNA repair and others in conjugation. The principal helicase in DNA replication is DnaB, which interacts with DnaG and other proteins to form the primosome. Topoisomerases - Bidirectional replication of the circular E. coli chromosome unwinds about 100,000 base pairs per minute. Relief of this torsional stress is essential for DNA replication to occur. Topoisomerases are enzymes with a swivel mechanism that can relieve this stress.")
16
Opptvinning av DNA med DnaB og SSB

17
The DnaB helicase

18
”Clamp loader”-komplekset monterer β-subenheten på DNA

19
Clamp og Clamp loader

20
Replikasjonsgaffel hos E. coli

21
DNA-replikasjon i E. coli: Trombonemodellen

23
DNA topoisomeraser endrer linking number (supercoiling)
")
24
Hvordan oppnås høy nøyaktighet?
Balansert nivå av de fire dNTP Polymerasereaksjonen har høy nøyaktighet 3’-5’-ekonukleaseaktiviteten til Pol I og Pol III fjerner feil som polymeraseaktiviteten innfører En rekke reparasjonssystemer tar seg av gjenværende feil

25
Korrekturlesning: DNA polymerase I fjerner et feilinkorporert nukleotid

26
5’-3’-eksonukleasedomenet i DNA polymerase I fjerner nukleotider i 5’ –enden av et nick

27
Krav til initiering: Aktivt initiator-protein (DnaA)
Supercoilet substrat - AT-rikt område

28
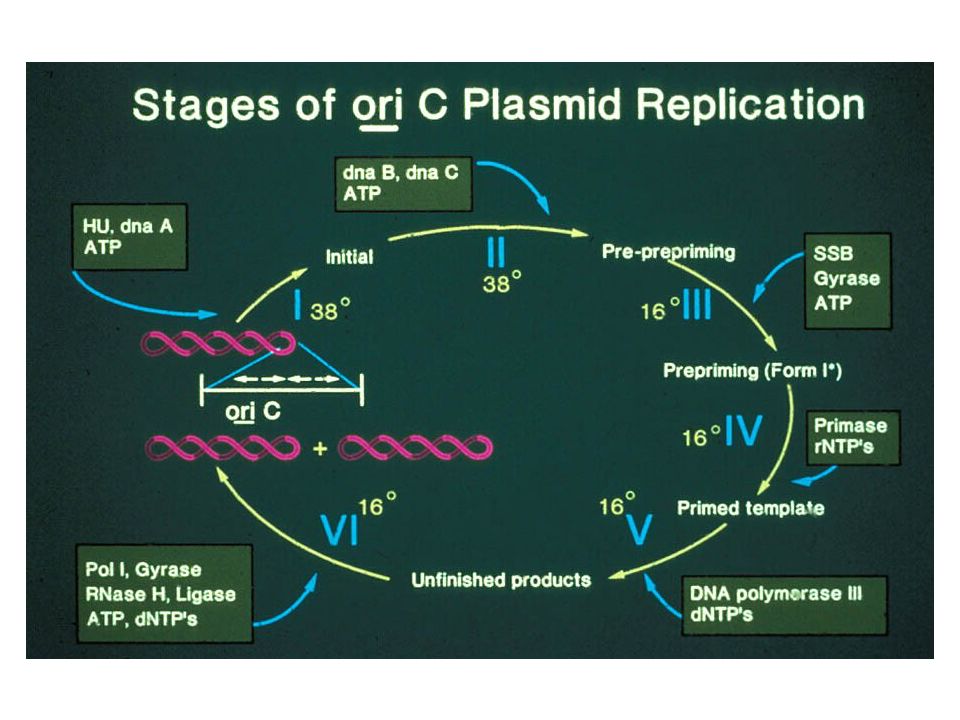
Replikasjonsorigo: oriC

30
The rate of DNA replication
E. coli: Genome, 4 Mb = 4 x 106 bp Fork rate approx bp / sec Replication time approx. 40 minutes = 2,400 secs Amount replicated by 2 forks in 40 mins = 2 x 2400 x 800 = 3,840,000 bp (~4 Mb)
")
31
Terminering av replikasjonen
TerG

32
DNA metylering I E. coli kan både cytosiner og adeniner bli enzymatisk metylert. Dcm metylasen lager 5-metylcytosin Dam metylasen lager 6-metyladenin Begge virker etter replikasjon. Cytosin metylering: Funksjon ukjent/restriksjon Adenin metylering er viktig for - Kontroll av DNA replikasjon - Reparasjon av DNA-skader - Genregulering - Beskyttelse mot fremmed DNA (?)
")
33
Identifikasjon av SeqA
oriC på et plasmid kan drive replikasjon Metylert oriC Dam+ : Metylert oriC Dam- : Umetylert oriC Dam- : Altså: Hemimetylert oriC blir ikke initiert. En mutant som mangler den negative regulator, vil tillate initiering av hemimetylert oriC, slik at Metylert oriC Dam- SeqA+:

34
Fullt metylert oriC Replikasjon gir hemimetylert oriC Elongering og separasjon Sekvestrering med SeqA (membran?) Remetylering Frigjøring

35
DnaA regulerer initieringstidspunktet.
Økende DnaA-konsentrasjon Tidligere initiering og ved lavere cellemasse (Løbner-Olesen et al, Cell 1989) Men er DnaA nødvendigvis REGULATOREN in vivo?
 Men er DnaA nødvendigvis REGULATOREN in vivo")
36
Three levels of negative control of DNA replication in E. coli:
1. Inactivation of the DnaA initiator 2. Sequestration of the origin of replication 3. Titration of DnaA to other chromosomal sites

37
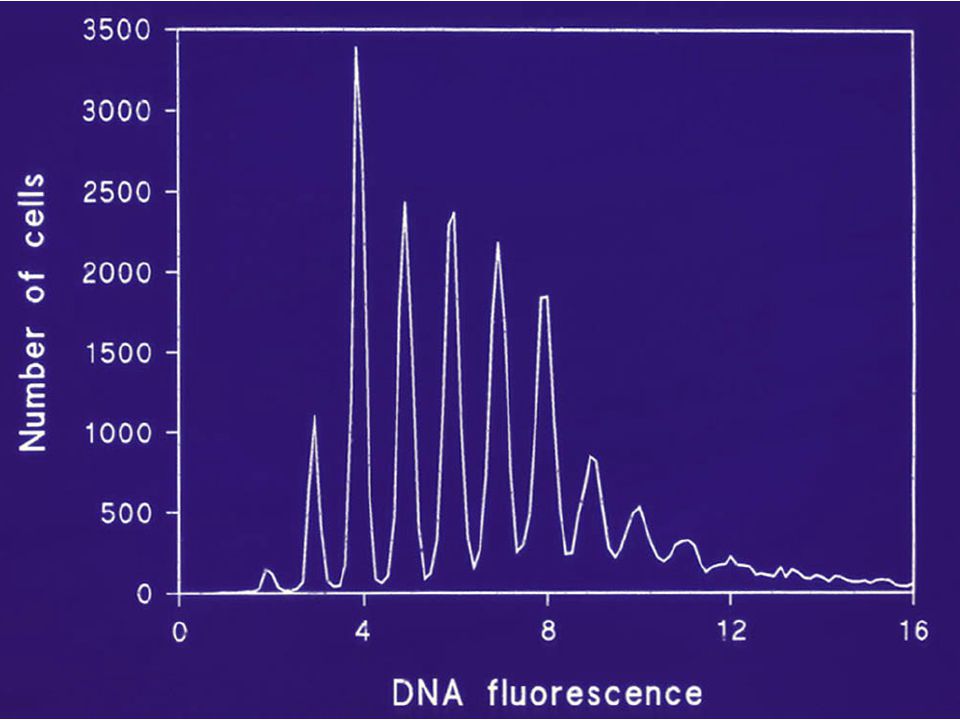
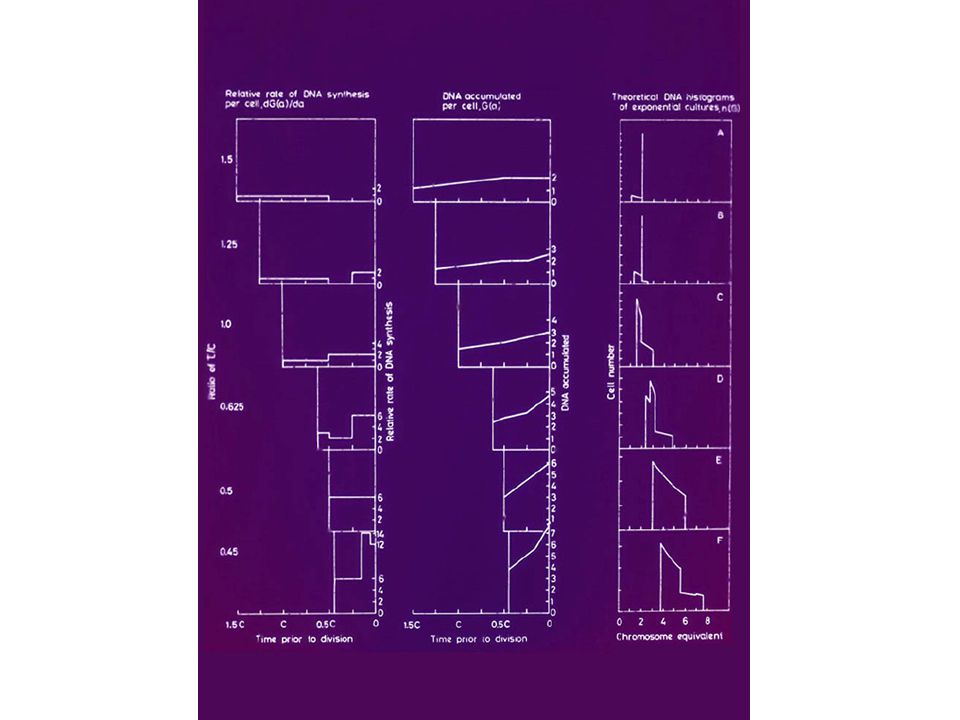
FLOW CYTOMETRY

40
Initiering av DNA replikasjon er avhengig av
Proteinsyntese RNA syntese Hemming av disse prosessene vil derfor gjøre at initiering stopper, mens elongering fortsetter. Sluttresultat: Antall hele kromosomer = antall oriC

42
Asynkroni: Når IKKE alle origins i en celle blir initiert samtidig

44
Mangel på initieringskontroll i en seqA mutant
50 min 4 4 Antall celler 30 min 8 8 20 min 8 8 Antall kromosomer (Lu et al, Cell 1994)
")
45
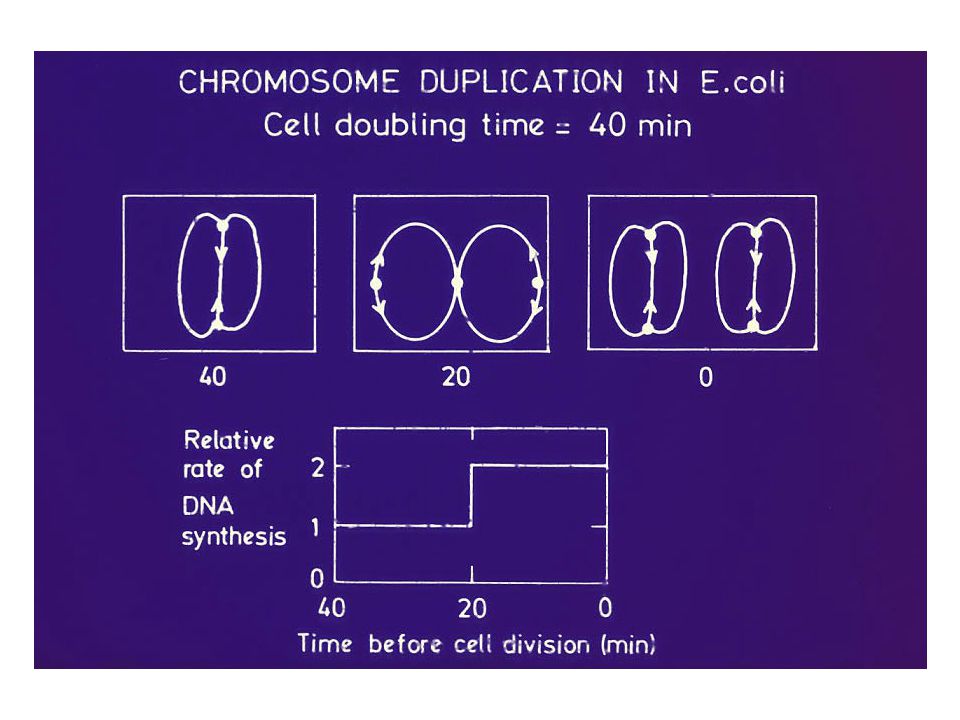
En generell cellesyklus i E. coli
Deling Deling Deling B C 40 min D 20 min C og D er konstante. Hvordan går det når doblingstiden blir under 60 min?

Liknende presentasjoner