Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
Forelesninger i BI 101 - Våren 2003 DE KJEMISKE BYGGESTEINER (Kap.3)
Tor-Henning Iversen, Plantebiosenteret (PBS), Institutt for biologi,NTNU Tlf PBS`s hjemmeside;
, Institutt for biologi,NTNU. Tlf PBS`s hjemmeside;")
2
Ansatte og hovedfagstudenter ved Plantebiosenteret i 2003
10 ansatte - vit.medarbeidere, teknisk personale, deltidsengasjerte forskere 5 dr. grads-stipendiater 6 hovedfagstudenter

3
Forelesninger BI 101 - vår 2003 GENERELL INFORMASJON
Lærebok : P.H. Raven & G.B. Johnson : Biology, 6th Edition (Mc Graw Hill) -2002 Sammendrag av de tre forelesninger som holdes av Tor-Henning Iversen – Kap. 3 De kjemiske byggesteiner (mandag 13. januar 2003) Kap. 8 Energi og metabolisme (onsdag 22. januar 2003) Kap. 10 Fotosyntese (torsdag 30. januar 2003) Vil bli lagt ut på fag-web-siden
 Sammendrag av de tre forelesninger som holdes av Tor-Henning Iversen – Kap. 3 De kjemiske byggesteiner (mandag 13. januar 2003) Kap. 8 Energi og metabolisme (onsdag 22. januar 2003) Kap. 10 Fotosyntese (torsdag 30. januar 2003) Vil bli lagt ut på fag-web-siden.")
4
Forelesninger i BI 101 - Våren 2003 MAKROMOLEKYLER (Kap
Forelesninger i BI Våren MAKROMOLEKYLER (Kap.3)- FUNKSJONELLE GRUPPER GENERELT: Selv om man har valgt å omtale de kjemiske byggesteiner dvs. biologiske makromolekyler i et eget kapittel så skal det huskes på at dette er basis for alle kapitler i læreboken. I dette kapittel fokuseres det på organiske molekyler dvs. kjemiske forbindelser som inneholder karbon (C). Hvis molekylet består av karbon og hydrogen (H) kalles det et hydrokarbon og er elektrisk nøytralt (upolart) Eksempel :propan-gass. Funksjonelle grupper er ladete (polare) og knyttet til hydrokarbon-kjernen (Figure 3.2). Kjemiske reaksjoner i en organisme vil normalt benytte funksjonelle grupper. Figure 3.2
- FUNKSJONELLE GRUPPER. GENERELT: Selv om man har valgt å omtale de kjemiske byggesteiner dvs. biologiske makromolekyler i et eget kapittel så skal det huskes på at dette er basis for alle kapitler i læreboken. I dette kapittel fokuseres det på organiske molekyler dvs. kjemiske forbindelser som inneholder karbon (C). Hvis molekylet består av karbon og hydrogen (H) kalles det et hydrokarbon og er elektrisk nøytralt (upolart) Eksempel :propan-gass. Funksjonelle grupper er ladete (polare) og knyttet til hydrokarbon-kjernen (Figure 3.2). Kjemiske reaksjoner i en organisme vil normalt benytte funksjonelle grupper. Figure 3.2.")
5
Forelesninger i BI 101 - Våren 2003 MAKROMOLEKYLER (Kap
Forelesninger i BI Våren MAKROMOLEKYLER (Kap.3)- HOVEDGRUPPER Biologiske makromolekyler (Table 3.1) er store komplekser med tusenvis av atomer og de har høye molekylvekter (> 1000) - f.eks. har hemoglobin en molekylvekt på og inneholder mer enn atomer. I levende celler finnes det 4 hovedgrupper av makromolekyler ; proteiner (bygget opp av aminosyrer) , nukleinsyrer (bygget opp av nukleotider), lipider og karbohydrater (bygget opp av sukker-monomerer som glukose), Sentralt i oppbyggingen av makromolekyler er dannelsen av kovalente bindinger gjennom en kondensasjons-reaksjon (dehydrering-syntese) hvor vann fjernes og energi tilføres (Figure 3.3a). Lipider er ikke er riktige makromolekyler da enkelt-byggesteinene i lipider ikke er kovalent bundet sammen. Hydrolyse (Figure 3.3b) er tilførsel av vann med samtidig frigjøring av energi ved at en kovalent binding brytes i makromolekylet.
- HOVEDGRUPPER. Biologiske makromolekyler (Table 3.1) er store komplekser med tusenvis av atomer og de har høye molekylvekter (> 1000) - f.eks. har hemoglobin en molekylvekt på og inneholder mer enn atomer. I levende celler finnes det 4 hovedgrupper av makromolekyler ; proteiner (bygget opp av aminosyrer) , nukleinsyrer (bygget opp av nukleotider), lipider og karbohydrater (bygget opp av sukker-monomerer som glukose), Sentralt i oppbyggingen av makromolekyler er dannelsen av kovalente bindinger gjennom en kondensasjons-reaksjon (dehydrering-syntese) hvor vann fjernes og energi tilføres (Figure 3.3a). Lipider er ikke er riktige makromolekyler da enkelt-byggesteinene i lipider ikke er kovalent bundet sammen. Hydrolyse (Figure 3.3b) er tilførsel av vann med samtidig frigjøring av energi ved at en kovalent binding brytes i makromolekylet.")
6
Forelesninger i BI 101 - Våren 2003 MAKROMOLEKYL - GRUPPER
Table 3.1

7
Forelesninger BI 101 - vår 2003 PROTEINER - ULIKE TYPER
Proteiner er bygget opp av et antall av maksimalt 20 ulike aminosyrer og foldet i rommet til en tredimensjonal form. De har en rekke oppgaver i cellen (se Table 3.2 i boken) ; enzym-katalyse, forsvars-mekanismer, transport, struktur-funksjon, bevegelser og regulering. Enzymer er mest kjent. Enzymer er globulære proteiner som er meget substrat-spesifikke og øker betydelig hastigheten på kjemiske reaksjoner - uten selv å bli omdannet. Ulike kjemiske grupper - prostetiske grupper (karbohydrater, lipider, fosfat- og hem-grupper m.fl.) - kan være bundet til proteinet. Forsvarsproteiner er lokalisert i cellemembranen som reseptorer og bruker sin form til å ”gjenkjenne” f.eks. bakterier og kreftceller Transportproteiner hjelper til å forflytte små molekyler og ioner. Hemoglobin transporterer oksygen i blodet og myoglobin oksygen i musklene. Strukturproteiner er fiber- eller trådlignende og finnes i form av keratin i hår, fibrin i blod, kollagen i hud m.fl. Bevegelsesproteiner finnes som protein-filamenter i muskler (actin og myosin) og som kontraktile proteiner i cellens cytoskjellett. Reguleringsproteiner finnes som hormoner og virker som intracellulære bærere i dyr. Reguleringsproteinene har også en regulatorisk funksjon f.eks. ved å slå på/slå av gener.
 ; enzym-katalyse, forsvars-mekanismer, transport, struktur-funksjon, bevegelser og regulering. Enzymer er mest kjent. Enzymer er globulære proteiner som er meget substrat-spesifikke og øker betydelig hastigheten på kjemiske reaksjoner - uten selv å bli omdannet. Ulike kjemiske grupper - prostetiske grupper (karbohydrater, lipider, fosfat- og hem-grupper m.fl.) - kan være bundet til proteinet. Forsvarsproteiner er lokalisert i cellemembranen som reseptorer og bruker sin form til å gjenkjenne f.eks. bakterier og kreftceller. Transportproteiner hjelper til å forflytte små molekyler og ioner. Hemoglobin transporterer oksygen i blodet og myoglobin oksygen i musklene. Strukturproteiner er fiber- eller trådlignende og finnes i form av keratin i hår, fibrin i blod, kollagen i hud m.fl. Bevegelsesproteiner finnes som protein-filamenter i muskler (actin og myosin) og som kontraktile proteiner i cellens cytoskjellett. Reguleringsproteiner finnes som hormoner og virker som intracellulære bærere i dyr. Reguleringsproteinene har også en regulatorisk funksjon f.eks. ved å slå på/slå av gener.")
8
Forelesninger BI 101 - vår 2003 OPPBYGGING AV AMINOSYRER
Aminosyrene er de grunnleggende byggesteiner i proteinet og 20 av disse inngår i proteiner. Hver aminosyre består av en aminogruppe, en karboksylgruppe, et hydrogen-atom og en sidekjede (R-gruppe) bundet til et sentralt C-atom. Sidekjeden er proteinets reaktive gruppe og viser mange ulike kjemiske egenskaper ( Figure 3.6) - noe som gir proteinet en bestemt funksjon og struktur. Av denne grunn klassifiseres aminosyrene avhengig av om sidekjedene er: ikke-polare (f.eks. leucin, 4 stk.), aminosyrer med polare men uladete sidekjeder (5 stk), ioniserbare dvs. elektrisk ladete (+ eller -: 5 stk),aromatiske (3 stk) og aminosyrer(prolin, methionin, cystein) med spesielle sidekjeder (3 stk). Cystein danner svovelbroer (-S-S-) med andre S-holdige aminosyrer.
 bundet til et sentralt C-atom. Sidekjeden er proteinets reaktive gruppe og viser mange ulike kjemiske egenskaper ( Figure 3.6) - noe som gir proteinet en bestemt funksjon og struktur. Av denne grunn klassifiseres aminosyrene avhengig av om sidekjedene er: ikke-polare (f.eks. leucin, 4 stk.), aminosyrer med polare men uladete sidekjeder (5 stk), ioniserbare dvs. elektrisk ladete (+ eller -: 5 stk),aromatiske (3 stk) og aminosyrer(prolin, methionin, cystein) med spesielle sidekjeder (3 stk). Cystein danner svovelbroer (-S-S-) med andre S-holdige aminosyrer.")
9
Forelesninger BI 101 - vår 2003 EKSEMPLER PÅ AMINOSYRER
Noen utvalgte aminosyrer - se også Figure 3.6

10
Forelesninger BI 101 - vår 2003 PEPTID-BINDING og POLYPEPTIDER
Proteiner er polymere av aminosyrer og gjennom en kondensasjonsreaksjon bindes to aminosyrer sammen ved dannelsen av en peptid-binding (se også Fig. 3.5 i boken). Kjeder av slike sammenkoblete aminosyrer kalles et polypeptid og en eller flere slike er med på å danne et protein. På en ende av polypeptidet er det en fri amino-gruppe ( N-(terminus)enden) ; på den andre enden er en fri karboksylgruppe (C- (terminus)enden).
. Kjeder av slike sammenkoblete aminosyrer kalles et polypeptid og en eller flere slike er med på å danne et protein. På en ende av polypeptidet er det en fri amino-gruppe ( N-(terminus)enden) ; på den andre enden er en fri karboksylgruppe (C- (terminus)enden).")
11
Forelesninger BI 101 - vår 2003 NIVÅER I PROTEINSTRUKTUREN
Figure 3.7 Proteinets fuksjon avhenger av formen på molekylet. Den tredimensjonale (3D) -formen kan undersøkes ved lavenergi eller røntgen (X)-analyser.Myoglobin var det første proteinet som ble analysert. Generelt kan sies at alle ”indre” aminosyrer er ikke-polare (leucin, valin, fenylalanin) dvs. det er lite ladete motsetninger internt og derfor tett mellom aminosyrene. Polare og ladete aminosyrer er på overflaten av proteinet. Protein-strukturen deles inn i ulike nivåer. Strukturen kan være enkel eller kompleks og inndeles i fire hoved nivåer (Fig. 3.7) ; primær, sekundær, tertiær og kvarternær samt i motifs(foldinger) og domains (funksjonelle enheter).
 -formen kan undersøkes ved lavenergi eller røntgen (X)-analyser.Myoglobin var det første proteinet som ble analysert. Generelt kan sies at alle indre aminosyrer er ikke-polare (leucin, valin, fenylalanin) dvs. det er lite ladete motsetninger internt og derfor tett mellom aminosyrene. Polare og ladete aminosyrer er på overflaten av proteinet. Protein-strukturen deles inn i ulike nivåer. Strukturen kan være enkel eller kompleks og inndeles i fire hoved nivåer (Fig. 3.7) ; primær, sekundær, tertiær og kvarternær samt i motifs(foldinger) og domains (funksjonelle enheter).")
12
Forelesninger BI 101 - vår 2003 NIVÅER I PROTEINSTRUKTUREN (forts.)
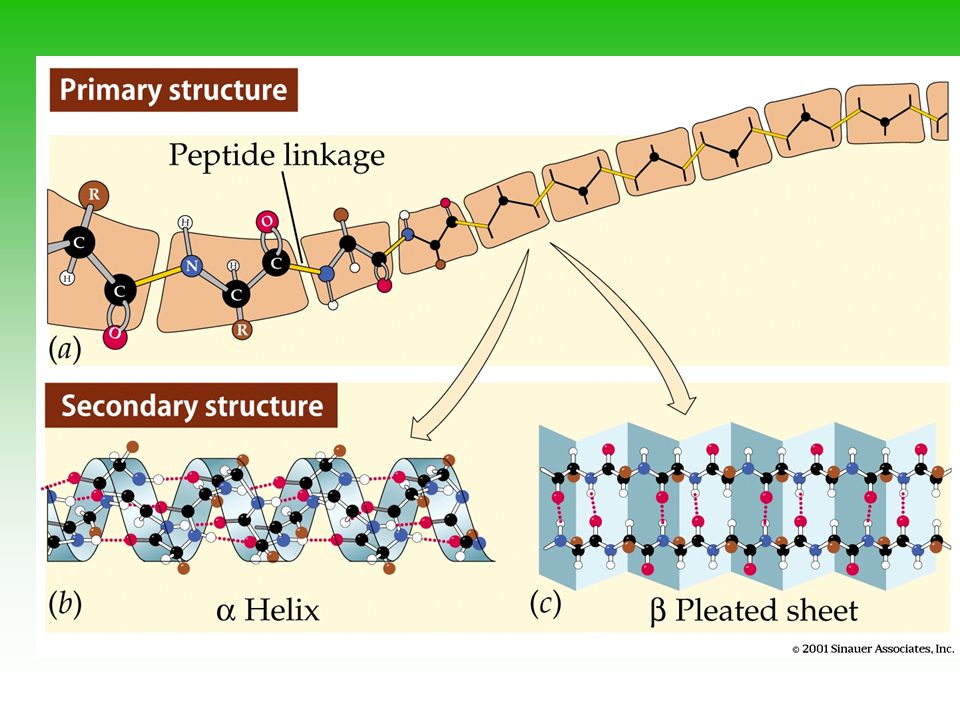
Primærstrukturen ; rekken av aminosyrer (diktert av et segment av et DNA-molekyl dvs. et gen - se senere). Oppdagelsen av sammenhengen mellom proteinets og DNA`s primærstruktur ga grunnlaget for moderne molekylærbiologi. Med 20 aminosyrer og ulike kombinasjonsmuligheter av disse blir det teoretiske antallet proteiner hvor man antar at proteinet består av 100 aminosyrer enormt; proteiner. Sekundærstrukturen ; oppbyggingen av denne krever hydrogen-bindinger mellom N-H i et peptid og C=O i et annet. Et eksempel er en -helix som er en høyredreiet spiral (neste bilde). Bindingene stabiliserer den dreiete formen - forutsatt at peptidene er bygget opp av korte sidekjeder. Mens -helix ofte finnes i struktur-fiberproteiner (f.eks. keratin i negler og hår), finnes en annen sekundærstruktur [-pleated sheet; (neste bilde)] i silke. Begge former finnes i enzymer. Motif ; Delene i sekundærstrukturen kan kombineres i proteiner på karakteristiske måter som kalles motif eller supersekundær-struktur. Et typisk eksempel er et ß ß-motif som danner en fold i proteinet. Eksempler (se boken) er ”Rossmann folden” , ß-barrel og turn motif.
. Oppdagelsen av sammenhengen mellom proteinets og DNA`s primærstruktur ga grunnlaget for moderne molekylærbiologi. Med 20 aminosyrer og ulike kombinasjonsmuligheter av disse blir det teoretiske antallet proteiner hvor man antar at proteinet består av 100 aminosyrer enormt; proteiner. Sekundærstrukturen ; oppbyggingen av denne krever hydrogen-bindinger mellom N-H i et peptid og C=O i et annet. Et eksempel er en -helix som er en høyredreiet spiral (neste bilde). Bindingene stabiliserer den dreiete formen - forutsatt at peptidene er bygget opp av korte sidekjeder. Mens -helix ofte finnes i struktur-fiberproteiner (f.eks. keratin i negler og hår), finnes en annen sekundærstruktur [-pleated sheet; (neste bilde)] i silke. Begge former finnes i enzymer. Motif ; Delene i sekundærstrukturen kan kombineres i proteiner på karakteristiske måter som kalles motif eller supersekundær-struktur. Et typisk eksempel er et ß ß-motif som danner en fold i proteinet. Eksempler (se boken) er Rossmann folden , ß-barrel og turn motif.")
14
Forelesninger BI 101 - vår 2003 NIVÅER I PROTEINSTRUKTUREN (forts.)
Tertiærstrukturen ; oppstår ved foldinger av polypeptidet (neste side) til en tredimensjonal form hvor ulike motifer bidrar. Stabiliteten av 3-D-formen avhenger av hvor godt den indre sammensetningen av aminosyrer fungerer sammen (les i boken om dette). Domains : Dette er en funksjonell enhet som er kodet av gen-sekvenser kalt exoner ( aminosyrer) og foldet som en uavhengig enhet. Ofte kan ulike domain ha spesifikke oppgaver (f.eks. binde en co-faktor eller enzym-substratet). Kvarternærstrukturen ; bygges opp ved at protein-underenheter i form av to eller flere polypeptider knyttes sammen for å danne et større molekyl (neste side). Som vist så er hemoglobin bygget av fire underenheter satt sammen ved ione- og hydrogen-bindinger.
 til en tredimensjonal form hvor ulike motifer bidrar. Stabiliteten av 3-D-formen avhenger av hvor godt den indre sammensetningen av aminosyrer fungerer sammen (les i boken om dette). Domains : Dette er en funksjonell enhet som er kodet av gen-sekvenser kalt exoner ( aminosyrer) og foldet som en uavhengig enhet. Ofte kan ulike domain ha spesifikke oppgaver (f.eks. binde en co-faktor eller enzym-substratet). Kvarternærstrukturen ; bygges opp ved at protein-underenheter i form av to eller flere polypeptider knyttes sammen for å danne et større molekyl (neste side). Som vist så er hemoglobin bygget av fire underenheter satt sammen ved ione- og hydrogen-bindinger.")
16
Hemoglobin (kvarternær struktur)
")
17
Forelesninger BI 101 - vår 2003 PROTEIN-DENATURERING OG BINDINGER
Både sekundær- og tertiær-strukturen er basert på primærstrukturen; ved forsiktig oppvarming av proteiner vil tertiær-strukturen forsvinne men ved avkjøling gjenvinnes formen kalt reversibel denaturering (se senere) Ved oppbyggingen av tertiærstrukturen gjennom interaksjoner mellom R-grupper, brukes andre bindingstyper enn H-bindinger ; S-broer, hydrofobe sidekjeder, van der Waals krefter og ionebindinger (saltbroer).
 Ved oppbyggingen av tertiærstrukturen gjennom interaksjoner mellom R-grupper, brukes andre bindingstyper enn H-bindinger ; S-broer, hydrofobe sidekjeder, van der Waals krefter og ionebindinger (saltbroer).")
18
Forelesninger BI 101 - vår 2003 PROTEINSTRUKTUREN - FOLDINGER
Hvordan foldes proteiner til en funksjonell form ? Foldinger av polypeptidet (se Figure 3.8) til en tredimensjonal form - var tidligere tenkt å skje spontant pga aminosyrenes ladninger. Men - foldingene er for kompliserte til å kunne skje uten cellens styring. Idag vet man at spesielle proteiner - chaperoniner - hjelper til for å få den riktige formen. Ialt er det påvist mer enn 17 ulike chaperoniner. Mange er varmesjokk-proteiner som produseres i store mengder hvis cellen utsettes for høy varme. Ved høy varme foldes proteiner ut (denatureres)- chaperoninene refolder (renaturerer) proteinene. Chaperoniner både sikrer riktig folding og ”redder” proteiner som er feil foldet. Det er antatt at mangel på chaperoniner fører til visse arvelige sykdommer f.eks. cystisk fibrose. Sykdommen antas å være forårsaket av en mutasjon i et gen som er ansvarlig for produksjonen av et transportprotein i cellemembranen. Proteinet har riktig aminosyrerekkefølge, men foldingene er mislykket. Det er også antatt at mangel på chaperoniner kan lede til Alzheimer.
 til en tredimensjonal form - var tidligere tenkt å skje spontant pga aminosyrenes ladninger. Men - foldingene er for kompliserte til å kunne skje uten cellens styring. Idag vet man at spesielle proteiner - chaperoniner - hjelper til for å få den riktige formen. Ialt er det påvist mer enn 17 ulike chaperoniner. Mange er varmesjokk-proteiner som produseres i store mengder hvis cellen utsettes for høy varme. Ved høy varme foldes proteiner ut (denatureres)- chaperoninene refolder (renaturerer) proteinene. Chaperoniner både sikrer riktig folding og redder proteiner som er feil foldet. Det er antatt at mangel på chaperoniner fører til visse arvelige sykdommer f.eks. cystisk fibrose. Sykdommen antas å være forårsaket av en mutasjon i et gen som er ansvarlig for produksjonen av et transportprotein i cellemembranen. Proteinet har riktig aminosyrerekkefølge, men foldingene er mislykket. Det er også antatt at mangel på chaperoniner kan lede til Alzheimer.")
19
Forelesninger BI 101 - vår 2003 PROTEINSTRUKTUREN - ENDRINGER
Hvordan utfoldes proteiner til en ikke-funksjonell form ? Utfoldinger av allerede foldete proteiner (se Figure 3.9) kalles denaturering. Dette kan skje ved endring i pH, temperatur eller ionestyrke i omgivelsene rundt proteinet. Denaturerte proteiner er biologisk inaktive. Det er spesielt viktig for enzymer at de er aktive proteiner. Enzymer i f.eks. magesekken er tilpasset ekstrem lav pH (pH2) og vil denaturere ved høyere pH, mens normale enzymer krever nøytral pH (pH7.4). Tilsvarende gjelder enzymer i organismer som lever under ekstreme forhold (varme kilder, lave temperaturer i arktiske områder) Må skille denaturering fra dissosiering av proteiner -f.eks. kan hemoglobin dissosiere i fire underenheter (to - og to -enheter) - uten at proteinet denatureres (Figure 3.10).
 kalles denaturering. Dette kan skje ved endring i pH, temperatur eller ionestyrke i omgivelsene rundt proteinet. Denaturerte proteiner er biologisk inaktive. Det er spesielt viktig for enzymer at de er aktive proteiner. Enzymer i f.eks. magesekken er tilpasset ekstrem lav pH (pH2) og vil denaturere ved høyere pH, mens normale enzymer krever nøytral pH (pH7.4). Tilsvarende gjelder enzymer i organismer som lever under ekstreme forhold (varme kilder, lave temperaturer i arktiske områder) Må skille denaturering fra dissosiering av proteiner -f.eks. kan hemoglobin dissosiere i fire underenheter (to - og to -enheter) - uten at proteinet denatureres (Figure 3.10).")
20
Forelesninger BI 101 - vår 2003 SLUTTKOMMENTARER OM PROTEINER
Overflaten og formen til proteinene bestemmer biologisk spesifisitet ved å gi muligheter for ikke-kovalente bindinger til andre molekyler (se figur). Dette er av stor betydning f.eks. på celleoverflaten (binder nærliggende celler, transport-mekanismer i cellemembranen, signal-mekanismer, antistoffdannelse m.m. - se senere forelesninger). Detaljert kunnskap om proteinformen og bindings-egenskapene kan anvendes f.eks. innen medisinsk behandling: eksempel er en protease som er viktig for dannelsen av nye HIV. Utviklingen av spesifikke inhibitorer som festes på overflaten av proteasen er viktig i å stoppe AIDS-utviklingen. Prioner (proteinaceous infectious particle) er en spesiell type proteiner som er selvreproduserende og årsak til mange ”nye” sykdommer - skrapesyke, kugalskap og Creutzfeldt-Jakobs sykdom
. Dette er av stor betydning f.eks. på celleoverflaten (binder nærliggende celler, transport-mekanismer i cellemembranen, signal-mekanismer, antistoffdannelse m.m. - se senere forelesninger). Detaljert kunnskap om proteinformen og bindings-egenskapene kan anvendes f.eks. innen medisinsk behandling: eksempel er en protease som er viktig for dannelsen av nye HIV. Utviklingen av spesifikke inhibitorer som festes på overflaten av proteasen er viktig i å stoppe AIDS-utviklingen. Prioner (proteinaceous infectious particle) er en spesiell type proteiner som er selvreproduserende og årsak til mange nye sykdommer - skrapesyke, kugalskap og Creutzfeldt-Jakobs sykdom.")
21
Forelesninger BI 101 - vår 2003 NUKLEINSYRER - INFORMASJONSMOLEKYLER
Nukleinsyrer er molekyler som lagrer, overfører og bruker informasjon. To typer arvemateriale; DNA (deoksyribo-nukleinsyre) og RNA (ribo-nukleinsyre). Koder for nedarving fra generasjon til generasjon i alle organismer gjennom dannelse av spesifikke aminosyresekvenser i proteiner. Informasjonstrømmen går fra DNA --> RNA. Med moderne mikroskopi er man idag istand til å se DNA-tråder. Utstyret - som kalles scanning-tunnel-mikroskopet - bruker elektroner og ikke lysstråler for å gjøre objektet synlig. I tillegg anvendes avansert datateknikk og prober for å skape en overflate av DNA-molekylet (se Figure 3.12).
 og RNA (ribo-nukleinsyre). Koder for nedarving fra generasjon til generasjon i alle organismer gjennom dannelse av spesifikke aminosyresekvenser i proteiner. Informasjonstrømmen går fra DNA --> RNA. Med moderne mikroskopi er man idag istand til å se DNA-tråder. Utstyret - som kalles scanning-tunnel-mikroskopet - bruker elektroner og ikke lysstråler for å gjøre objektet synlig. I tillegg anvendes avansert datateknikk og prober for å skape en overflate av DNA-molekylet (se Figure 3.12).")
22
Forelesninger BI 101 - vår 2003 NUKLEINSYRENES STRUKTUR
Nukleinsyrene er lange polymerer bygget opp av monomerene nukleotider som består av tre komponenter : pentose-sukker, en fosfat-gruppe og en organisk nitrogen-base (purin eller pyrimidin) - se Fig Hvis fosfat-gruppen mangler kalles molekylet et nukleosid. Nukleotidene bindes sammen med kovalente bindinger - fosfo-diester-bindinger - mellom sukker i ett og fosfat i neste nukleotid (Fig. 3.14).
 - se Fig Hvis fosfat-gruppen mangler kalles molekylet et nukleosid. Nukleotidene bindes sammen med kovalente bindinger - fosfo-diester-bindinger - mellom sukker i ett og fosfat i neste nukleotid (Fig. 3.14).")
23
Forelesninger BI 101 - vår 2003 MER OM DNA
Organismer koder gjennom strukturen av DNA for hvordan proteiner skal bygges opp. Dette skjer ved å spesifisere informasjon om hvordan rekkefølgen av aminosyrer skal være (genetisk kode). Kodene er ulike kombinasjoner av fire nukleotider i spesifikke sekvenser - CGTTACG..... DNA eksisterer i cellen (kjernen og noe i mitokondrier/kloroplaster) som dobbeltkjeder (dobbel-helix). Opp-bygging kan sammenlignes med en vindeltrappe. Hvert trinn er et basepar hvor én base i en kjede er bundet med H-bindinger til en komplementær base i den andre. Adenin(A) vil alltid parres med thymin (T) - cytosin (C) med guanin (G). Merk 5`- og 3`-enden som er viktig ved påbygningen av kjedene.
. Kodene er ulike kombinasjoner av fire nukleotider i spesifikke sekvenser - CGTTACG..... DNA eksisterer i cellen (kjernen og noe i mitokondrier/kloroplaster) som dobbeltkjeder (dobbel-helix). Opp-bygging kan sammenlignes med en vindeltrappe. Hvert trinn er et basepar hvor én base i en kjede er bundet med H-bindinger til en komplementær base i den andre. Adenin(A) vil alltid parres med thymin (T) - cytosin (C) med guanin (G). Merk 5`- og 3`-enden som er viktig ved påbygningen av kjedene.")
24
Forelesninger BI 101 - vår 2003 DNA SAMMENLIGNET MED RNA
RNA skiller seg fra DNA på tre måter : 1. Inneholder ribose-sukker og ikke deoksyribose , 2. Har uracil istedet for thymin og 3. Er vanligvis enkelttrådet pga mangel på enzymer som kan etablere en dobbel-helix. RNA kopieres (tran-skriberes) fra DNA som en enkelttråd (Figure 3.16). Det finnes flere RNA-former som deltar i syntesen av proteiner - m-RNA, r-RNA og t-RNA Figure 3.16
 fra DNA som en enkelttråd (Figure 3.16). Det finnes flere RNA-former som deltar i syntesen av proteiner - m-RNA, r-RNA og t-RNA. Figure")
25
Forelesninger BI 101 - vår 2003 Trenger vi et ”Gen-Krim” ?
Kronikk i Aftenposten ; mange positive reaksjoner for at et slikt behov bør dekkes Oppgaver ; påvise fremmed DNA i planter som innføres til Norge fungere som en ”karantene-stasjon” - forutprøving av transgene planter Utredning om et nasjonalt kontroll-laboratorium for GMP i Norge ; konkrete planer om å legge laboratoriet til Plantebiosenteret

26
Forelesninger BI 101 - vår 2003 LIPIDER
Lipider omfatter en gruppe makromolekyler som er løst definert men med det fellestrekk at de er uløselige i vann. Felles-egenskapen - deres vann-uløselighet - er av sentral betydning for lipiders funksjon i transport av ioner og molekyler gjennom cellemembraner. De vanligste lipider er fett og oljer (som lagrer energi) og fosfolipider (som er bærekonstruksjonen i membraner). Ulike typer organismer inneholder andre typer fett enn lipider; Terpener er langkjedete lipider og inngår i pigmenter (klorofyll dvs. lysabsorberende pigmenter, synspigmentet retinal). Steroider som består av fire C-ringer (f.eks. kolesterol, testosteron, østrogen og signalmolekyler) Prostaglandiner er en gruppe på 20 lipider som er modifiserte fettsyrer med to upolare haler bundet til en 5-C-ring. Virker som lokale kjemiske budbærere i vev i pattedyr.
 og fosfolipider (som er bærekonstruksjonen i membraner). Ulike typer organismer inneholder andre typer fett enn lipider; Terpener er langkjedete lipider og inngår i pigmenter (klorofyll dvs. lysabsorberende pigmenter, synspigmentet retinal). Steroider som består av fire C-ringer (f.eks. kolesterol, testosteron, østrogen og signalmolekyler) Prostaglandiner er en gruppe på 20 lipider som er modifiserte fettsyrer med to upolare haler bundet til en 5-C-ring. Virker som lokale kjemiske budbærere i vev i pattedyr.")
27
Forelesninger BI 101 - vår 2003 FOSFOLIPIDER DANNER MEMBRANER
Fosfolipider er kjernen i biologiske membraner, har ett polart ”hode” og to lange upolare ”haler”. Den polare delen er vanntiltrekkende (hydrofil) ; halene er vannavstøtende (hydrofobe) (Figure 3.18). Figure 3.18
 ; halene er vannavstøtende (hydrofobe) (Figure 3.18). Figure")
28
Forelesninger BI 101 - vår 2003 FOSFOLIPIDER DANNER MEMBRANER
Phosphatidyl-cholin er et eksempel på et fosfolipid Fosfolipidet generelt består av tre typer underenheter : Glyserol : en tre-C-alkohol hvor hvert C-atom bærer en OH-gruppe. Bærebjelken i fosfolipidene. Fettsyrer : lange kjeder av C-H bindinger (hydrokarbon-kjeder) som ender i en COOH-gruppe. To fettsyrer er forankret i ett glyserol-molekyl. Fosfat-gruppe : er bundet til den ene enden av glyserol. Har ofte et ladet molekyl bundet til seg (cholin, ethanolamin, eller aminosyren serin)
 som ender i en COOH-gruppe. To fettsyrer er forankret i ett glyserol-molekyl. Fosfat-gruppe : er bundet til den ene enden av glyserol. Har ofte et ladet molekyl bundet til seg (cholin, ethanolamin, eller aminosyren serin)")
29
Forelesninger BI 101 - vår 2003 ANDRE TYPER LIPIDER
Fett og oljer er enkle lipider. Fett består av et glyserol-molekyl med tre fettsyrer påkoblet - derav fellesnavnet triglycerid (Fig. 3.19) . Mer korrekt er navnet triacylglycerol. Fett er i fast form mens oljer er flytende ved romtemperatur. Må lære den grunnleggende oppbygging av triglyceridene. Siden triglyceridene mangler en polar ende er de hydrofobe. I vann klumper de spontant sammen og danner fettkuler.
 . Mer korrekt er navnet triacylglycerol. Fett er i fast form mens oljer er flytende ved romtemperatur. Må lære den grunnleggende oppbygging av triglyceridene. Siden triglyceridene mangler en polar ende er de hydrofobe. I vann klumper de spontant sammen og danner fettkuler.")
30
Forelesninger BI 101 - vår 2003 METTEDE OG UMETTEDE FETTSYRER
Mettede fettsyrer har enkle bindinger mellom C-atomene i H-C-kjeden dvs. alle bindinger er mettet med H-atomer (f.eks. palmintinsyre; Fig.3-20a) - umettede har en eller flere dobbeltbindinger - se f.eks. Fig. 3.20b som viser linolje-syre (18 C-atomer). De mettede har rette kjeder – de umettede får en knekk ved dobbelbindingen. Fettsyrer med flere dobbeltbindinger kalles flerumettede.
 - umettede har en eller flere dobbeltbindinger - se f.eks. Fig. 3.20b som viser linolje-syre (18 C-atomer). De mettede har rette kjeder – de umettede får en knekk ved dobbelbindingen. Fettsyrer med flere dobbeltbindinger kalles flerumettede.")
31
Forelesninger BI 101 - vår 2003 FETT SOM NÆRING
Fett inneholder 40 C-atomer og forholdet mellom C og C-H-bindinger er avgjørende for energien som kan hentes ut fra fettet. Sammenlignet med karbohydrater er fettmolekylene mye mer effektive pga den store mengden C-H-bindinger mht lagring av energi. I gjennomsnitt gir fett 9 kcal kjemisk energi pr. gram - karbohydrater 4 kcal/g. Med unntak av visse fiskeoljer så er animalsk fett mettet mens plantefett (untatt kokosolje og palmeolje) er umettet. Oljer kan overføres til fast fett ved å tilføre hydrogen. Generelt så er umettet fett helsemessig bedre enn mettet. En diett med høyt fettinnhold øker sjansen for hjerte-/karsykdommer som følge av dannelsen av fettavleiringer i blodkarene. En hovedårsak til hjerneslag og hjerteinfarkt skyldes fettpropper som brytes løs ett sted i kroppen og forflyttes til hjerte/hjerne hvor en blokkering skjer slik at oksygen ikke når frem.
 er umettet. Oljer kan overføres til fast fett ved å tilføre hydrogen. Generelt så er umettet fett helsemessig bedre enn mettet. En diett med høyt fettinnhold øker sjansen for hjerte-/karsykdommer som følge av dannelsen av fettavleiringer i blodkarene. En hovedårsak til hjerneslag og hjerteinfarkt skyldes fettpropper som brytes løs ett sted i kroppen og forflyttes til hjerte/hjerne hvor en blokkering skjer slik at oksygen ikke når frem.")
32
Forelesninger BI 101 - vår 2003 KARBOHYDRATER -MONOSAKKARIDER
Karbohydrater er ulike typer forbindelser som stort sett har felles generell empirisk formel (CH2O)n - (n er antallet av C-atomer). Følgende hovedgrupper finnes; monosakkarider, disakkarider, oligosakkarider og polysakkarider. Monosakkarider : Glukose [C6H12O6 eller (CH2O)6 ] som har to former; rette kjeder (1%) og ringer i vandige omgivelser (99%), er det dominerende monosakkarid i alle celler. Dannes i planter ved fotosyntesen og brukes av alle celler som energi-kilde hvor glukosen omdannes til vann og karbondioksid pluss energi. Pentoser er 5-karbon sukkere som finnes bl.a. i cellveggen hos planter. Forøvrig er ribose og deoksyribose de viktigste som byggesteiner i RNA og DNA. Figure 3.21 6-carbon sugars
n - (n er antallet av C-atomer). Følgende hovedgrupper finnes; monosakkarider, disakkarider, oligosakkarider og polysakkarider. Monosakkarider : Glukose [C6H12O6 eller (CH2O)6 ] som har to former; rette kjeder (1%) og ringer i vandige omgivelser (99%), er det dominerende monosakkarid i alle celler. Dannes i planter ved fotosyntesen og brukes av alle celler som energi-kilde hvor glukosen omdannes til vann og karbondioksid pluss energi. Pentoser er 5-karbon sukkere som finnes bl.a. i cellveggen hos planter. Forøvrig er ribose og deoksyribose de viktigste som byggesteiner i RNA og DNA. Figure carbon sugars.")
33
Forelesninger BI 101 - vår 2003 DISAKKARIDER og POLYSAKKARIDER
Ved kondensasjons-reaksjoner og dannelsen av kovalente glykosidiske bindinger kobles to monosakkarider sammen til et disakkarid (sukrose, laktose (Figure 3.23) cellobiose, maltose, m.fl.). Maltose og cellobiose dannes begge fra to molekyler glukose, men ved respektivt en - og -1,4 glykosidisk binding. Polysakkarider : Dette er makromolekyler bygget opp av monosakkarid-underenheter. Stivelse og cellulose er begge bygget fra glukose og hovedgruppene av polysakkarider i planter; glykogen finnes i dyr (se senere) Figure 3.23
 cellobiose, maltose, m.fl.). Maltose og cellobiose dannes begge fra to molekyler glukose, men ved respektivt en - og -1,4 glykosidisk binding. Polysakkarider : Dette er makromolekyler bygget opp av monosakkarid-underenheter. Stivelse og cellulose er begge bygget fra glukose og hovedgruppene av polysakkarider i planter; glykogen finnes i dyr (se senere) Figure")
34
Forelesninger BI 101 - vår 2003 SUKKER-ISOMERE
Glukose er ikke det eneste sukkeret som har den empiriske formelen C6H12O6.Andre vanlig forekommende 6-C-sukkere med samme formel er fruktose og galaktose. Dette er isomere eller alternative former av glukose. Men - atomene er arrangert på ulik måte dvs. deres tredimensjonale-form er ulik. Også smaken på slike strukturelle isomere (Figure 3.24) er forskjellig - fruktose smaker søtere enn glukose. De to sukkere danner også ulike polymere. Stereoisomere sukkere (Figure 3.24) som f.eks. galaktose og glukose, har samme bindingstruktur, men ulik orientering på én OH-gruppe. Disse gruppene er speilbilder av hverandre. Selv en så liten forskjell har dramatisk effekt på dannelsen av polymere. Figure 3.24
 er forskjellig - fruktose smaker søtere enn glukose. De to sukkere danner også ulike polymere. Stereoisomere sukkere (Figure 3.24) som f.eks. galaktose og glukose, har samme bindingstruktur, men ulik orientering på én OH-gruppe. Disse gruppene er speilbilder av hverandre. Selv en så liten forskjell har dramatisk effekt på dannelsen av polymere. Figure")
35
Forelesninger BI 101 - vår 2003 SAMMENBINDING AV SUKKERE
Transport av sukkere i kroppen skjer som glukose dvs. et monosakkarid i blodet. I planter skjer dette i form av disakkarider (oftest sukrose =glukose + fruktose) som er mer stabil mot uønsket nedbrytning. Åraken er at det ikke finnes disakkarid-nedbrytende enzymer i alle celler, men kun i de hvor glukose trengs og disakkaridet nedbrytes til glukose. Oligosakkarider bygges opp av flere monosakkarider som er bundet sammen ved glykosidiske bindinger på flere steder. I tillegg har de ofte tilknyttet funksjonelle grupper som gir dem spesielle egenskaper. I plasmamembranen stikker det ofte ut av cellen oligosakkarider bundet til proteiner (kalt glykoproteiner) og lipider (kalt glykolipider) som virker som celle-gjenkjenning-signaler .
 som er mer stabil mot uønsket nedbrytning. Åraken er at det ikke finnes disakkarid-nedbrytende enzymer i alle celler, men kun i de hvor glukose trengs og disakkaridet nedbrytes til glukose. Oligosakkarider bygges opp av flere monosakkarider som er bundet sammen ved glykosidiske bindinger på flere steder. I tillegg har de ofte tilknyttet funksjonelle grupper som gir dem spesielle egenskaper. I plasmamembranen stikker det ofte ut av cellen oligosakkarider bundet til proteiner (kalt glykoproteiner) og lipider (kalt glykolipider) som virker som celle-gjenkjenning-signaler .")
36
Forelesninger BI 101 - vår 2003 LAGRINGS-POLYSAKKARIDER
Organismene lagrer metabolsk energi i monosakkarider ved å overføre dem til disakkarider (f.eks. maltose = glukose + glukose). Maltose-molekyler bindes sammen i lange kjeder til uløselige former av polysakkarider ; stivelse og cellulose (se Figure 3.27) som lagres i spesifikke organer i planter (f.eks. i poteter og ved ). Hos dyr finnes glykogen som er et energi-lager i lever og muskler. Mens stivelse lett lar seg bryte ned av enzymer (bl.a. amylase i spytt) er cellulose i celleveggen meget lite nedbrytbar. Av stivelse er det to former ; amylose og amylopektin. Figure 3.27
. Maltose-molekyler bindes sammen i lange kjeder til uløselige former av polysakkarider ; stivelse og cellulose (se Figure 3.27) som lagres i spesifikke organer i planter (f.eks. i poteter og ved ). Hos dyr finnes glykogen som er et energi-lager i lever og muskler. Mens stivelse lett lar seg bryte ned av enzymer (bl.a. amylase i spytt) er cellulose i celleveggen meget lite nedbrytbar. Av stivelse er det to former ; amylose og amylopektin. Figure")
37
Forelesninger BI 101 - vår 2003 STRUKTUR -POLYSAKKARIDER
Graden av forgreninger i glykosidiske bindinger representerer forskjellen mellom cellulose (u-forgrenet), stivelse og glykogen (mest forgrenet) – (se Figure 3.26 og 3.27). Cellulose er - i motsetning til stivelse som består av -glukose - bygget opp av - glukose. Cellulose er et struktur-polysakkarid og hovedkomponenten i alle plantecellevegger. Drøvtyggere kan utnytte celleulose som næring da de har en tarmflora av bakterier og protister som har cellulose-nedbrytende enzymer (cellulaser).
, stivelse og glykogen (mest forgrenet) – (se Figure 3.26 og 3.27). Cellulose er - i motsetning til stivelse som består av -glukose - bygget opp av - glukose. Cellulose er et struktur-polysakkarid og hovedkomponenten i alle plantecellevegger. Drøvtyggere kan utnytte celleulose som næring da de har en tarmflora av bakterier og protister som har cellulose-nedbrytende enzymer (cellulaser).")
38
Forelesninger BI 101 - vår 2003 STRUKTUR -POLYSAKKARIDER
Omdannete ( derivative) karbohydrater er kjemisk endret ved at de er påhengt funksjonelle grupper ( f.eks. fosfat-, amino- og N-acetyl-grupper). Hovedgrupper av slike kalles sukkerfosfater (a; viktig i energiomsetning), aminosukkere (b; i brusk/bindevev) og kitin [skjellettstruktur i insekter, krabber, hummer (Figure 3.28) og sopp]. Kitin er en modifisert form av cellulose hvor en N-gruppe er knyttet til glukose-enheten. Sammen med proteiner danner den et tett, ugjennomtrengelig skall som kun få organismer kan nedbryte. Figure 3.28
 karbohydrater er kjemisk endret ved at de er påhengt funksjonelle grupper ( f.eks. fosfat-, amino- og N-acetyl-grupper). Hovedgrupper av slike kalles sukkerfosfater (a; viktig i energiomsetning), aminosukkere (b; i brusk/bindevev) og kitin [skjellettstruktur i insekter, krabber, hummer (Figure 3.28) og sopp]. Kitin er en modifisert form av cellulose hvor en N-gruppe er knyttet til glukose-enheten. Sammen med proteiner danner den et tett, ugjennomtrengelig skall som kun få organismer kan nedbryte. Figure")
Liknende presentasjoner