Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
Regulering av DNA Transkripsjon i Eukaryote Organismer
ID, Kull 99, Vår 2001 Frank Skorpen IKM, DMF

2
Regulering av gen-uttrykk på mange nivåer
Klargjøring av DNA Transkripsjon Initiering Stopp hnRNA prosessering mRNA Transport Translasjon Stabilitet Protein Kjemisk modifisering Prosessering Lokalisering Stabilitet

3
Gen-uttrykk reguleres primært ved regulering av transkripsjons-initiering
Klargjøring av DNA Transkripsjon Initiering Stopp hnRNA prosessering mRNA Transport Translasjon Stabilitet Protein Kjemisk modifisering Prosessering Lokalisering Stabilitet

4
Nukleære RNA polymeraser
RNA pol I transkriberer rRNA gener RNA pol II transkriberer protein-kodende gener (klasse II gener) RNA pol III transkriberer tRNA gener
 RNA pol III. transkriberer tRNA gener.")
5
Geners regulatoriske områder
TRANSKRIPSJONSENHET 5’ 3’ Exon Intron TATAAT konsensus: Start TATA Inr -200 -50 -25 +1 Proximal Distal ENHANCER ELEMENTER CORE PROMOTER

6
Mulige transkripsjonsfaktor-bindende elementer kan identifiseres ut i fra DNA sekvens
E2F

7
Forskjellige sett av aktivatorer aktiverer forskjellige gener
B C Gen 1 A D E Gen 2

8
Forskjellige typer celler uttrykker forskjellige gener fordi de inneholder forskjellige sett av transkripsjons-aktivatorer Celletype I Celletype II B D E B A D E C A A A C

9
Transkripsjons-aktivatorer rekrutterer transkripsjonsmaskineriet til promoteren
Aktiverings- domene TATA Inr DNA-bindings- domene -25 +1 CORE PROMOTER

10
Mange aktivatorer binder som homo-
eller heterodimérer Dimériserings- domene TATA Inr -200 -50 Proximal Distal -25 +1 ENHANCER ELEMENTER CORE PROMOTER

11
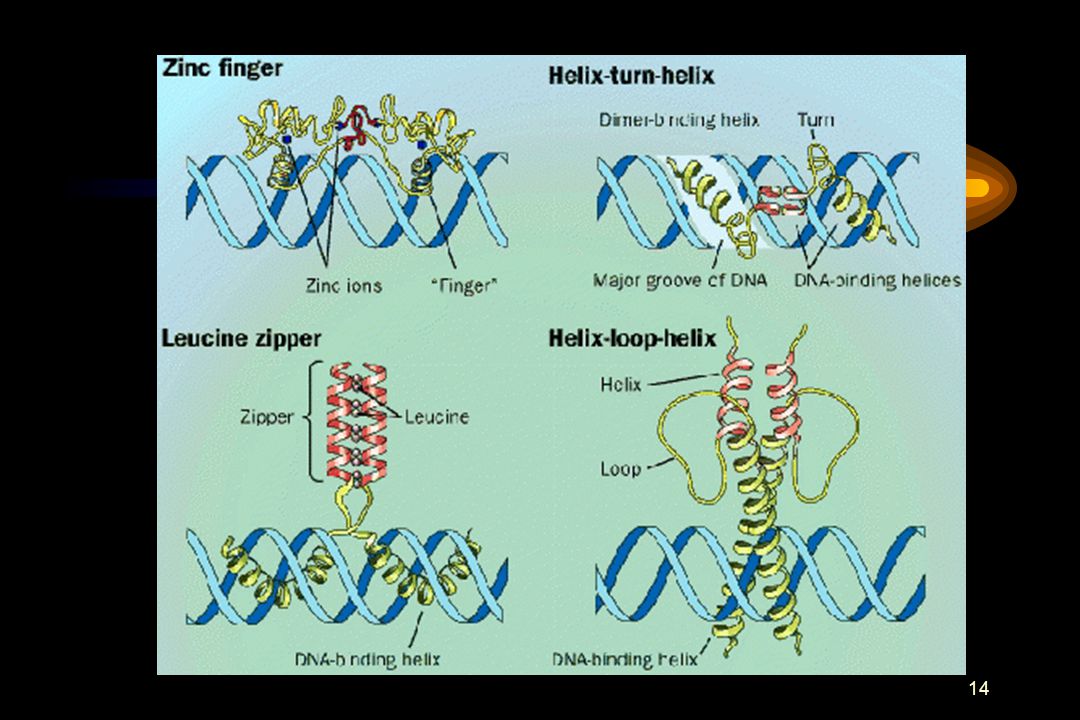
Transkripsjons-aktivatorer: familier
Helix-turn-helix Leucine zipper Helix-loop-helix Zinc finger andre

12
Helix-turn-helix Leucine zipper + NH2 COOH HOOC
Leucine zipper proteiner fungerer som dimérer av to subenheter, hver delt i to helix-regioner. Leucine (L) residier i C- terminalen danner en hydrofob overflate for interaksjon mellom de to protein sub- enhetene. Positive aminosyrer N-terminalt for leucinene danner kontaktflaten med DNA. Helix-turn-helix 2 3 COOH 1 NH2 Helix-turn-helix motivet finnes i en rekke prokaryote regulatoriske proteiner og i homeodomain proteiner i eukaryoter. Helix 3 er “recognition helix” som kontakter major groove i DNA helixen. Helix 1 and 2 ligger over helix 3 og kan kontakte andre proteiner.
 residier i C- terminalen danner en hydrofob overflate. for interaksjon mellom de to protein sub- enhetene. Positive aminosyrer N-terminalt. for leucinene danner kontaktflaten med DNA. Helix-turn-helix COOH. 1. NH2. Helix-turn-helix motivet finnes i en rekke. prokaryote regulatoriske proteiner og i. homeodomain proteiner i eukaryoter. Helix 3. er recognition helix som kontakter major. groove i DNA helixen. Helix 1 and 2 ligger over helix 3 og kan. kontakte andre proteiner.")
13
Helix-loop-helix C2H2 zinc finger + Zn
Diagram av en zinc finger bestående av to cysteine (C) og to histidine (H) residier koordinert rundt et sink atom. Andre høyt konserverte aminosyrer i denne klassen av zinc fingre er også vist. Zinc fingre kan også bruke fire cysteiner til å koordinere zinc atomet, slik som i steroid hormon receptorer. + NH2 HOOC COOH Helix Loop P C L H Zn Y COOH C2H2 zinc finger NH2 Helix-loop-helix proteiner har to helixer bundet sammen med en loop av ukjent struktur. Disse helixene danner overflaten for dannelsen av dimérer. En tredje helix rik på positive aminosyrer er ansvarlig for DNA-binding.
 og to histidine (H) residier. koordinert rundt et sink atom. Andre høyt. konserverte aminosyrer i denne klassen av. zinc fingre er også vist. Zinc fingre kan også. bruke fire cysteiner til å koordinere zinc. atomet, slik som i steroid hormon receptorer. + NH2. HOOC. COOH. Helix. Loop. P. C. L. H. Zn. Y. COOH. C2H2 zinc finger. NH2. Helix-loop-helix proteiner har to helixer. bundet sammen med en loop av ukjent. struktur. Disse helixene danner overflaten. for dannelsen av dimérer. En tredje helix. rik på positive aminosyrer er ansvarlig. for DNA-binding.")
15
Transkripsjons-aktivatorer kan slås på og av
PROTEIN- SYNTESE LIGAND- BINDING FOSFORYLERING BINDING TIL ANNEN SUBENHET INAKTIV P DNA-bindende subenhet AKTIV Aktiverende subenhet

16
Signal-aktivering av gener
Steroid hormon reseptor Steroid hormon TRANSKRIPSJON P P Insulin Signal- kaskade Insulin reseptor

17
Aktivatorer rekrutterer RNA pol II og et sett av 5 generelle transkripsjonsfaktorer
IIF IIH IIE IIB IID Aktivator TATA Inr -50 +1

18
TBP binder til TATA-element og danner en plattform for binding av de andre faktorene
TAFIIs TBP Activator TATA Inr -50 +1

19
Rekruttering skjer vanligvis i to trinn: først TFIID,
deretter resten av komplekset IIE IIH RNA pol II IIF IIB IID Aktivator TATA Inr -50 +1

20
Generelle transkripsjonsfaktorer og RNA pol II responderer ikke til aktivatorer in vitro
IIF IIH IIE IIB IID Aktivator TATA Inr -50 +1

21
Aktivatorer virker vanligvis gjennom ikke-DNA-bundne ko-aktivatorer
RNA pol II IIF IIH IIE IIB TBP The Core Srb/Mediator Complex Srb2, -4, -5, -6, and -7; (pos) Med1, -2, -4, -6, -7, and -8; (pos) Gal11, Sin4, Rgr1, Rox3, and Pgd1 (neg) Srb4 Srb10 CDK (neg) Srb11 Srb9 Activator TATA Inr -50 +1
 Med1, -2, -4, -6, -7, and -8; (pos) Gal11, Sin4, Rgr1, Rox3, and Pgd1 (neg) Srb4. Srb10 CDK. (neg) Srb11. Srb9. Activator. TATA. Inr")
22
In vivo foreligger DNA i form av kromatin. Kromatin virker hemmende på
DNA transkripsjon. H2B H2A H3 H4 H1 DNA helix Nukleosom: bp DNA tvunnet rundt en oktamer av histoner

23
Histon acetyltransferaser (HATs) og histon deacetylaser (HDACs)
H2B H2A H3 H4 + H20 CH3 C=O OH S Coenzyme A SH

24
HAT enzymer kan dirigeres til promotere ved å binde til aktivatorer
CREB c-Jun Fos P/CAF p300/CBP P/CAF: “p300/CBP-Associating Factor” p300/CBP, “CBP: CREB-binding protein” E2F-1 p53 Nukleosom

25
TAFII250 har intern HAT aktivitet
TFIID TAFII250 HAT aktivitet Aktivator TATA

26
Mange repressorer fungerer ved å rekruttere HDAC aktiviteter
E2F/DP Rb HDAC1 Deacetylering og represjon Mad Max SIN3 YY1 HDAC2

27
SWI/SNF bruker ATP til å remodellere nukleosomstrukturer
Swi - Snf Complex ATP ADP + Pi Swi2 Aktivator +1 TATA Inr

28
SWI/SNF kan rekrutteres som del av
RNA pol II holoenzym Swi - Snf Complex ATP ADP + Pi Swi2 RNA pol II holoenzym Aktivator +1 TATA Inr

29
Rekruttering ved synergi
RNA pol II IIF IIH IIE IIB Swi - Snf ATP ADP + Pi HATs Srb/Mediator TAFII250 TBP Aktivator +1 TATA Inr DNA-bøyende ko-aktivator

30
Modell av pre-initieringskompleks
Aktivatorer Generelle (basale) transkripsjonsfaktorer Ko-aktivatorer/ TBP-assosierte faktorer RNA pol II
 transkripsjonsfaktorer. Ko-aktivatorer/ TBP-assosierte faktorer. RNA pol II.")
31
Produksjon av mRNA Exon Intron 5’ 3’ Kopiering til pre-mRNA
AAAAAAA-3’ Fjerning av introns, påsetting av m7G(5’)pp cap, 3’-ende-kløyving og polyadenylering 5’-m7Gpp mRNA Exon Intron
pp cap, 3’-ende-kløyving og polyadenylering. 5’-m7Gpp. mRNA. Exon. Intron.")
32
elongating RNA pol II (Pol IIo)
RNA pol II holoensym er involvert i både RNA-kjede forlengelse, RNA prosessering, og terminering av transkripsjon Fjerning av intron elongating RNA pol II (Pol IIo) pre mRNA AAUAAA Kløyvings- / polyadenylerings- faktorer Elongerings- Spleise- CTD (YSPTSPS)n
 pre. mRNA. AAUAAA. Kløyvings- / polyadenylerings- faktorer. Elongerings- Spleise- CTD. (YSPTSPS)n.")
33
Oppsummering: Transkripsjonsaktiviteten av et gen bestemmes av hvor effektivt RNA pol II holoenzym rekrutteres til promoteren Dette bestemmes av den synergistiske effekten av de til enhver tid bundne aktivatorer Gener reguleres forskjellig fordi de binder forskjellige sett av aktivatorer Aktivatorer virker vanligvis gjennom ikke-DNA-bundne ko-aktivatorer

34
In vivo foreligger DNA i form av kromatin
In vivo foreligger DNA i form av kromatin. Kromatin hemmer transkripsjons-initiering Assosiert med RNA pol II finnes enzymaktiviteter som re-modellerer kromatin, slik som SWI/SNF og HATs/HDACs SWI/SNF og HATs/HDACs kan også rekrutteres via interaksjon med sekvens-spesifikke aktivatorer Forskjellige promotere har forskjellige krav til sammensetningen av RNA pol II holoenzym

Liknende presentasjoner