Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
Hvordan få oversikten?

2
Annotering av sekvensen

3
Kromosom 16: et av de minste

4
Finding genes What are we looking for? Where are we looking?
Proteins encoded in mRNA Non-coding RNA (ncRNA) genes Where are we looking? Prokaryotes Eukaryotes (often introns)
 genes. Where are we looking Prokaryotes. Eukaryotes (often introns)")
5
Source: Eddy SR (2001) Nature Reviews in Genetics
Classes of RNA fRNA: Functional RNA — essentially synonymous with non-coding RNA mRNA: Messenger RNA — coding for proteins miRNA: MicroRNA — putative translational regulatory gene family ncRNA: Non-coding RNA — all RNAs other than mRNA rRNA: Ribosomal RNA siRNA: Small interfering RNA — active molecules in RNA interference snRNA: Small nuclear RNA — includes spliceosomal RNAs snmRNA: Small non-mRNA — essentially synonymous with small ncRNAs snoRNA: Small nucleolar RNA — usually involved in rRNA modification stRNA: Small temporal RNA — e.g. lin-4 and let-7 in C. elegans tRNA: Transfer RNA Source: Eddy SR (2001) Nature Reviews in Genetics
 Nature Reviews in Genetics.")
6
Informasjon i sekvensen som kan brukes for å finne gener
”Signaler” i sekvensen: Spleisesignaler, promotere, termineringssignaler, polyA-signaler, CpG-øyer (Gene search by signal) ”Innholdet” i sekvensen: ORFs, kodonstatistikk osv.(Gene search by content) Likhet med kjente gener (Gene search by similarity)
 Innholdet i sekvensen: ORFs, kodonstatistikk osv.(Gene search by content) Likhet med kjente gener (Gene search by similarity)")
7
Fra gen til protein: så lett for cellen, så vanskelig for oss

9
Simple protein finding
Examine all 6 possible reading frames 3 frames on forward strand 3 frame on reverse strand Plot positions of Initiation (start) (Methionine) codon: ATG Termination (stop) codons: TAA, TAG, TGA Look for long stretches without stop codons after a start codon Source:
 (Methionine) codon: ATG. Termination (stop) codons: TAA, TAG, TGA. Look for long stretches without stop codons after a start codon. Source:")
10
Standard Genetic Code The standard genetic code is used in most organisms Another code is use din mitochondria and some organisms Overview of gentic codes in various organisms:

11
Start and stop codon distribution
Distribution of start codons (short lines) and stop codons (long lines) in the six reading frames along a genomic sequence (lacZ operon in E.coli) There is an open reading frame (lacZ) in frame +3 from position 1284 to 4355. Created by DNA STRIDER.
 and stop codons (long lines) in the six reading frames along a genomic sequence (lacZ operon in E.coli) There is an open reading frame (lacZ) in frame +3 from position 1284 to Created by DNA STRIDER.")
12
Prokaryotic promotor regions
Source:

13
Transcription termination

14
Shine-Dalgarno (SD) sequence
The 16S rRNA ribosomal protein binding site

16
Transcription and translation
Genomic DNA Promotor Terminator Exon1 Intron1 Exon2 Intron2 Exon3 Primary transcript GU…AG GU…AG 3’UTR 5’UTR Spliced mRNA AAAA… Cap Start AUG Stop TAA/TAG/TGA Protein M

17
Gene, exon and intron number for whole ExInt and subdivisions
Gene number Exon number Intron number Whole ExInt 94 615 Non-redundant ExInt 15 271 Rattus norvegicus 835 4889 7191 Homo sapiens 8287 60 499 43 127 Mus musculus 3044 18 920 15 407 Drosophila melanogaster 15 220 64 271 89 969 Caenorhabditis elegans 18 924 Arabidopsis thaliana 25 216 Saccharomyces cerevisiae 589 1695 1438

18
Fordeling av eksonstørrelser i ExInt

19
Fordeling av intronstørrelser i ExInt

20
Intron-fase: ekson/intron-overganger mellom kodoner eller i dem
Intron phase 1 2 All ExInt (49%) (28%) (23%) Non-redundant (48%) (28%) (24%) Rattus norvegicus 2842 (39%) 2365 (33%) 1384 (28%) Mus musculus 6703 (44%) 5921 (38%) 2783 (18%) Caenorhabditis elegans (47%) (26%) (27%) Homo sapiens (44%) (36%) 8602 (20%) Arabidopsis thaliana (56%) (22%) (22%) Drosophila melanogaster (42%) (32%) (26%) Saccharomyces cerevisiae 641 (45%) 428 (30%) 369 (25%)
 (28%) (23%) Non-redundant (48%) (28%) (24%) Rattus norvegicus (39%) 2365 (33%) 1384 (28%) Mus musculus (44%) 5921 (38%) 2783 (18%) Caenorhabditis elegans (47%) (26%) (27%) Homo sapiens (44%) (36%) 8602 (20%) Arabidopsis thaliana (56%) (22%) (22%) Drosophila melanogaster (42%) (32%) (26%) Saccharomyces cerevisiae. 641 (45%) 428 (30%) 369 (25%)")
21
Hvordan finne spleisesignaler og eksoner?
Vektsmatriser: Hvordan er fordelingen av nukleotider rundt spleiseseter? ”Weight array matrices” hvor det tas hensyn til nabonukleotider ”Maximal dependence decomposition”: Korrelasjoner med ikke-nabonukleotider Skjulte Markov-modeller Neurale nettverk: En mønstergjenkjenningsteknikk som ”lærer”

22
Slik lages en vektmatrise

23
Og slik brukes den

24
Konsensus-sekvenser for ekson/intron-overganger

25
Forskjellige klasser av eksoner som må oppdages på forskjellige måter
Innledende eksoner: Begynner med et startkodon og slutter med et spleisedonorsete Interne eksoner: Begynner med et akseptorsete og slutter med et donorsete Terminale eksoner: Begynner med et akseptorsete og slutter med et stoppkodon Enkelteksongener: Begynner med et startkodon og slutter med et stoppkodon

27
Integrert genfinning: Hva følger etter hva?
Ved å integrere resultater fra f. Eks signalsøk og innholdssøk kan nøyaktigheten av teknikkene forbedres

28
Neuronnettverk: et eksempel
with a positive value and others with a negative value; sums these values; and then converts them to an output of approximately 0 or 1. The system is trained using a set of known coding sequences, and as each sequence is utilized, the strengths and types of connections (positive or negative) between the neurons are adjusted, decreasing or increasing the signal to the next neuron in a manner that produces the correct output. The major difference between neural networks for exon and secondary structure prediction is that the exon prediction uses sequence pattern information as input whereas secondary structure prediction uses a window of amino acid sequence in the protein. In Grail II, a candidate sequence is evaluated by calculating pattern frequencies in the sequence and applying these values to the neural network. If the output is close to a value of 1, then the region is predicted to be an exon. The Grail II system for finding exons in eukaryotic genes (Uberbacher and Mural 1991; Uberbacher et al. 1996). The method uses a neural network to identify patterns characteristic of coding sequences. The network includes three layers, an input layer for the data with the data coming from a candidate exon sequence, and a hidden layer for discerning relationships among the input data. An output layer comprising one neuron indicates whether or not the region is likely to be an exon. Each neuron receives information from a set in the layer above, some
 between the neurons are adjusted, decreasing or increasing the signal to the next neuron in a manner that produces the correct output. The major difference between neural networks for exon and secondary structure prediction is that the exon prediction uses sequence pattern information as input whereas secondary structure prediction uses a window of amino acid sequence in the protein. In Grail II, a candidate sequence is evaluated by calculating pattern frequencies in the sequence and applying these values to the neural network. If the output is close to a value of 1, then the region is predicted to be an exon. The Grail II system for finding exons in eukaryotic genes (Uberbacher and Mural 1991; Uberbacher et al. 1996). The method uses a neural network to identify patterns characteristic of coding sequences. The network includes three layers, an input layer for the data with the data coming from a candidate exon sequence, and a hidden layer for discerning relationships among the input data. An output layer comprising one neuron indicates whether or not the region is likely to be an exon. Each neuron receives information from a set in the layer above, some.")
29
Sekvens”innhold”: Forskjeller mellom den ekte leserammen og de to andre
Ramme 1 er den ekte, og inneholder kodoner som koder for et protein med gjennomsnittlig aminosyresammensetning

30
Kodonbruk i de tre leserammene
Dersom kodonpreferenser tas i betraktning vil slike tabeller bli avhengige av organismen

31
Basefordeling på de tre kodonposisjonene

32
Å skille mellom kodende og ikkekodende sekvenser ut fra basesammensetningen av de tre kodonposisjonene Antall ganger en base forekommer i hver av de tre kodonposisjonene i vinduet = Nij. Forventet verdi for hver base i hver av de tre kodonposisjonene Eij=(Ni1+Ni2+Ni3)/3 Divergensen D=Σ|Eij-Nij| Vindu: 67 kodoner EMBL-databasen 1984
/3. Divergensen D=Σ|Eij-Nij| Vindu: 67 kodoner EMBL-databasen")
33
Codon usage in the E.coli genome
Source:

34
Codon usage in the human genome
Source:

35
Codon usage diagram Usage of various codons along the sequence of lacZ
O: Optimal codon usage S: Suboptimal codon usage R: Rare codon usage U: Unique codon usage Created by DNA STRIDER.

36
Comparative genomics methods
Gene finding by sequence comparison to sequences known to be transcribed or translated Compare the genomic sequence to sequence databases Proteins mRNA sequences EST sequences (mRNA) Both exact matches and approximate matches are interesting Conserved sequences between species Program: Procrustes
 Both exact matches and approximate matches are interesting. Conserved sequences between species. Program: Procrustes.")
38
Et eksempel på et resultat med søkeprogrammet Genscan

39
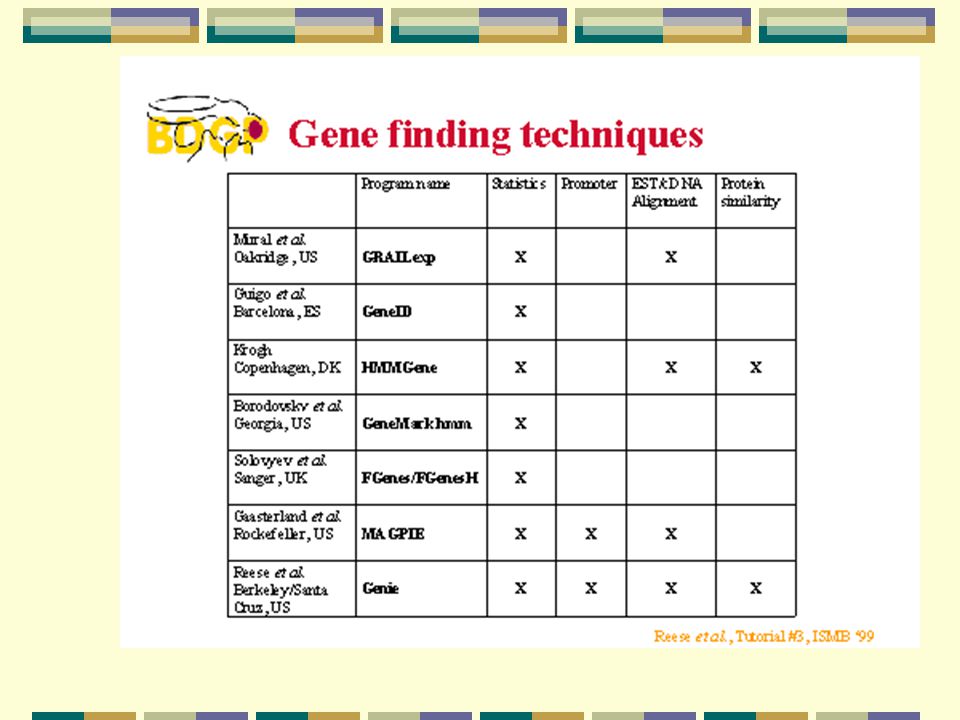
Genfinnere på nettet

40
Så lett for cellen, så vanskelig for oss

42
Skjulte Markov-modeller (HMM): Et enkelt eksempel
For genfinning vil tilstandene (states) være eksoner, introner og evt andre ønskede sekvensklasser (5’ og 3’ UTR, promoterområder, intergeniske områder, repetitivt DNA osv.). Transisjonssannsynlighetene vil variere med tilstanden (intron kan bare følges av internt eller terminalt ekson osv). Sannsynligheten for overgang fra ekson til intron avhenger av lokal sekvens, bare høy ved plausible spleiseseter
 være eksoner, introner og evt andre ønskede sekvensklasser (5’ og 3’ UTR, promoterområder, intergeniske områder, repetitivt DNA osv.). Transisjonssannsynlighetene vil variere med tilstanden (intron kan bare følges av internt eller terminalt ekson osv). Sannsynligheten for overgang fra ekson til intron avhenger av lokal sekvens, bare høy ved plausible spleiseseter.")
Liknende presentasjoner


















>")





