Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
Oversikt RNA polymerase I RNA polymerase III RNA polymerase II
proteinfaktorer og promoter/gen-organisering RNA polymerase III RNA polymerase II Kromatin som medspiller 80% av total RNA syntese fra disse ”Oddpols”

2
RNA polymerase I (RNAPI)
")
3
Klasse I gener (rRNA) transkribert av RNA polymerase I (RNAPI)
RNAPI syntetiserer kun en type RNA: ribosomalt RNA En transkripsjonsenhet (7.5 kB) Multiple tandem-gener for å øke rRNA-produksjonen genkopier avhengig av art
 Multiple tandem-gener for å øke rRNA-produksjonen genkopier avhengig av art.")
4
Nucleoli RNAPI lokalisert til nucleoli (ribosom-fabrikker)
")
5
Klasse I gener (rRNA): promoter-organisering
Repeterte gener med promotere imellom Intergenisk spacer (IGS) med terminator+promoter Core UPE Terminator elementer Enhancere
 med terminator+promoter. Core. UPE. Terminator. elementer. Enhancere.")
6
Klasse I gener (rRNA): Hjelpefaktorer involvert
Repeterte gener med promotere imellom Intergenisk spacer (IGS) med terminator+promoter UBF UBF SL1 TTF-1 RNAPI Core UPE Terminator elementer Enhancere
 med terminator+promoter. UBF. UBF. SL1. TTF-1. RNAPI. Core. UPE. Terminator. elementer. Enhancere.")
7
RNA polymerase III (RNAPIII)
")
8
Klasse III-gener transkribert av RNA polymerase III
RNAPIII syntetiserer et fåtall ulike RNA som er små, stabile og ikke-translaterte tRNA, 5S rRNA, 7SL RNA, U6 snRNA ++

9
Promotere - tre typer Typer promotere
Type I: som i 5S rRNA genet, A-I-C blokker Type II: som i tRNA genene, A+B blokker Type III: atypiske uten intrageniske elementer A I C A B

10
Klasse III gener transkribert av RNA polymerase III
Promotere med intrageniske elementer Mutasjoner endrer både promoter og produkt Polymerasen må lese igjennom aktiverende transkripsjonskomplekser A-blokk B-blokk

11
Klasse III gener: hjelpefaktorer som deltar
Type II promotere - mest klassiske Sekvens: TFIIIC - TFIIIB/TBP - RNAPIII RNAPIII TFIIIC A B TFIIIB TBP

12
Klasse III gener: hjelpefaktorer som deltar
Type I promotere en type gen, en spesial faktor TFIIIA Sekvens: TFIIIA - TFIIIC - TFIIIB/TBP - RNAPIII RNAPIII TFIIIC TFIIIA A I C TFIIIB TBP

13
RNA polymerase II (RNAPII)
")
14
Understanding transcription - Increasing complexity
80ties 70ties Figure 1. Fundamental elements of eukaryotic transcriptional control. (A) Early studies led to the identification of multiple eukaryotic RNA polymerases and the mapping of promoter and enhancer DNA sequences. Core promoter elements (grey) that direct transcription by RNA Pol II (yellow, green shading) include the TATA box (TATA), the initiator (INR), and the downstream promoter element (DPE). Promoter-proximal basal level enhancer elements (BLE, GC box, CCAAT box, green) were found in many genes; however, gene-specific signal-responsive distal enhancer elements were also identified and are represented here as the hormone responsive element (HRE, purple) and nuclear factor element (NFE, blue). (B) Sequence-specific DNA-binding transcription factors that were isolated biochemically include specificity protein-1 (Sp1, green), families of activator proteins (i.e., AP-1, green), CCAAT enhancer-binding proteins (C/EBP), steroid receptors (i.e., glucocorticoid or GR, purple) and tissue-specific transcription factors (i.e., NF- B, blue). Components of the core initiation machinery required by RNA Pol II for promoter recognition and basal transcription from naked DNA were also fractionated biochemically, including TATA-binding protein, (TBP, red) and general transcription factors (TFIIA, TFIIB, TFIID, TFIIE, TFIIF and TFIIH, purple). (C) Because TBP is insufficient for activated transcription, discovery of TAF subunits (orange) within TFIID revealed a requirement for co-activators to mediate activator responsiveness. Depicted here is one example of a specific and functional interaction between Sp1 and Drosophila TAF110 (human TAF130). (D) Many co-activators and co-repressors were subsequently found to be required for mediating signals between sequence-specific transcription factors and the core machinery. For further discussion pertaining to the specific co-regulators depicted, see text and Table 1. 90ties Today Lemon and Tjian 2000 Genes Dev. 14:
 Early studies led to the identification of multiple eukaryotic RNA polymerases and the mapping of promoter and enhancer DNA sequences. Core promoter elements (grey) that direct transcription by RNA Pol II (yellow, green shading) include the TATA box (TATA), the initiator (INR), and the downstream promoter element (DPE). Promoter-proximal basal level enhancer elements (BLE, GC box, CCAAT box, green) were found in many genes; however, gene-specific signal-responsive distal enhancer elements were also identified and are represented here as the hormone responsive element (HRE, purple) and nuclear factor element (NFE, blue). (B) Sequence-specific DNA-binding transcription factors that were isolated biochemically include specificity protein-1 (Sp1, green), families of activator proteins (i.e., AP-1, green), CCAAT enhancer-binding proteins (C/EBP), steroid receptors (i.e., glucocorticoid or GR, purple) and tissue-specific transcription factors (i.e., NF- B, blue). Components of the core initiation machinery required by RNA Pol II for promoter recognition and basal transcription from naked DNA were also fractionated biochemically, including TATA-binding protein, (TBP, red) and general transcription factors (TFIIA, TFIIB, TFIID, TFIIE, TFIIF and TFIIH, purple). (C) Because TBP is insufficient for activated transcription, discovery of TAF subunits (orange) within TFIID revealed a requirement for co-activators to mediate activator responsiveness. Depicted here is one example of a specific and functional interaction between Sp1 and Drosophila TAF110 (human TAF130). (D) Many co-activators and co-repressors were subsequently found to be required for mediating signals between sequence-specific transcription factors and the core machinery. For further discussion pertaining to the specific co-regulators depicted, see text and Table 1. 90ties. Today. Lemon and Tjian Genes Dev. 14:")
15
Klasse II-gener transkribert av RNA polymerase II
Mediator Signal RD RNAPII TF TAD TFIID DBD Transcription factors TBP Chromatin GTFs TATA Enhancer Promoter Nucleosomal template - chromatin modifying activities

16
Språket: cis-elementene

17
cis-elementenes funksjon = Templater for kompleksdannelse
The function of cis-elements is being templates for the assembly of multiprotein complexes

18
Promoter- organisering
core promoter oppstrøms regulatoriske element Enhancere Boundary elements LCR - locus control regions Promoter- organisering

19
Klasse II gener transkribert av RNA polymerase II
Subklasser av RNAPII-promotere mRNA-kodende TATA+ INR+ både TATA og INR uten TATA, uten INR snRNA-kodende

20
Noen typer cis-elementer
UPE - Upstream promoter elementer Binder konstitutivt uttrykte faktorer som finnes i alle celler Lokalisert nær TATA/INR (innen ca. 200 bp) Eksempler CCAAT boks - binder ulike TFs (CTF/NF-I, CBFINF-Y) GC-rike bokser - binder Sp1 Regulatoriske elementer 1. Responsive elementer eks.: CRE, HSE, GRE - medierer respons overfor cAMP, heat shock, glucocorticoider 2. Celletypespesifikke elementer Lokalisert innblandet med UPE
 Eksempler. CCAAT boks - binder ulike TFs (CTF/NF-I, CBFINF-Y) GC-rike bokser - binder Sp1. Regulatoriske elementer. 1. Responsive elementer. eks.: CRE, HSE, GRE - medierer respons overfor cAMP, heat shock, glucocorticoider. 2. Celletypespesifikke elementer. Lokalisert innblandet med UPE.")
21
Enhancere Distale elementer - Enhancere
Øker kraftig aktiviteten av en promoter virker over lang avstand, uavh. av orientering, upstream/downstream Drosophila wing margin enhancer: 85 kb upstream TSS Immunoglobulin Hm enhancer: i 2. Intron T-cell receptor a-chain enhancer: 69 kb downstream samme cis-elementer som proksimalt multiple cis-elementer innen et lite område (50 bp kb) cis-elementer for mange ulike faktorer responsive/vevsspesifikke avhengig av sammensetning
 cis-elementer for mange ulike faktorer. responsive/vevsspesifikke avhengig av sammensetning.")
22
Klasse II gener transkribert av RNA polymerase II
TATA TFIID TBP TF RNAPII Mediator GTFs Chromatin Signal

23
RNA polymerase II Core med aktivt sete RPB1 (´-like)binder DNA
Prokaryot RNA polymerase II ´ Eukaryot DNA-bindende Core med aktivt sete RPB1 (´-like)binder DNA RPB2 (-like) binder NTP RPB3 (-like) assembly factor Felles subenheter RPB5, 6 og 8 felles for RNAPI-III Til forskjell fra prokaryot RNAP, er RNAPII ikke i stand til spesifikk promoter-gjenkjenning NTP-binding
binder DNA. RPB2 (-like) binder NTP. RPB3 (-like) assembly factor. Felles subenheter. RPB5, 6 og 8 felles for RNAPI-III. Til forskjell fra prokaryot RNAP, er RNAPII ikke i stand til spesifikk promoter-gjenkjenning. NTP-binding.")
24
CTD - ”C-terminal domain”
Hale på største subenhet: (YSPTSPS)n n = 26 i gjær, 52 human pol.II hydrofil eksponert hale Unik for RNAPII Essensiell funksjon in vivo >50% letalt
n. n = 26 i gjær, 52 human pol.II. hydrofil eksponert hale. Unik for RNAPII. Essensiell funksjon in vivo. >50% letalt.")
25
CTD- funksjon Fosforylering: (YSPTSPS)n
Reversibel fosforylering på både Ser og Tyr Fosforylering endres gjennom transkripsjonssyklus Fosforylering skjer etter PIC assembly defosforylering av fri pol eller ved terminering

26
Fosforylerings-syklus

27
CTD funksjon Initiering Promoter clearance Elongering
Rolle i “rekruttering” av RNAPII til promoter bare ufosforylert pol deltar i PIC-assembly Promoter clearance RNAPII går over i hyperfosforylert elongeringsmodus CTD fosforylering bryter interaksjoner og RNAPII frikobles fra PIC Elongering Samtidig: CTD fosforylering danner nye interaksjoner med elongeringsfaktorer En rekke nye CTD-bindende proteiner identifisert siste år med funksjoner i splicing og terminering Tett kobling: transkripsjon - pre-mRNA prosessering

28
CTD binder også en rekke faktorer
Holoenzym = Mediator + core RNAPII Mediator RNA processering RNAPII = ”mRNA factory” som utfører koblet transkripsjon, capping, splicing og prosessering av 3´-ende

29
Klasse II gener transkribert av RNA polymerase II
TATA TFIID TBP TF RNAPII Mediator GTFs Chromatin Signal

30
GTFs - generelle transkripsjonsfaktorer
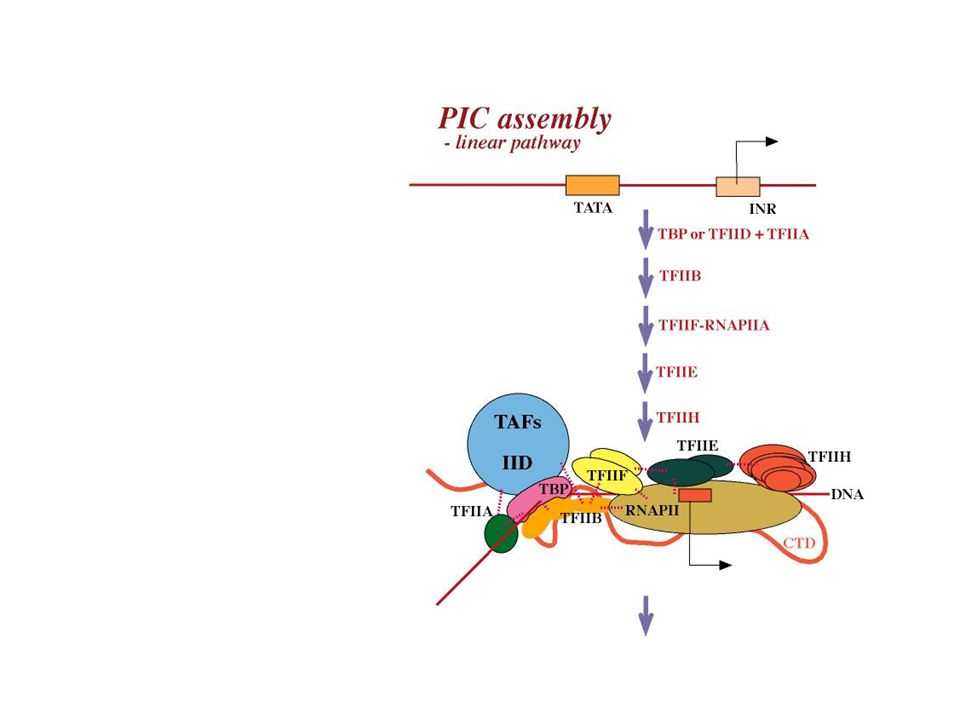
Hjelpefaktorer som dirigerer RNAPII til promoter Trinnvis oppbygging Dannelse av PIC - pre-initiation complex

32
1. Trinn: TATA bindes av TBP
TBP binder TATA - sekvensgjenkjenning minor groove kontakt TBP binder også en rekke andre polypeptider aktivatorer TAFs (kalles da TFIID = TBP + TAFs) GTFs (TFIIB, TFIIA) TBP = universell TF involvert i alle 3 tr.systemene TBP i SL1, TFIID, TFIIIB protein DNA
 GTFs (TFIIB, TFIIA) TBP = universell TF involvert i alle 3 tr.systemene. TBP i SL1, TFIID, TFIIIB. protein. DNA.")
33
TBP = sadel (med TATA som hest)
3D: Sadel-struktur Konkav innerside binder DNA i minor groove via 10-stranded antiparallelt b-sheet Konveks overside binder prot via 4 a-helikser stigbøyler (“stirrup”) på hver ende
 på hver ende.")
34
TBP bøyer DNA (stakkars hest)
Ikke slik, …….men slik

35
RNA polymerasen gjenkjenner en GTF-bundet promoter
TBP TFIIA TFIIB Dette protein-promoter komplekset er det som gjenkjennes av RNAPII

36
PIC ”modnes” etter RNAPII- binding
TFIIH: multisubenhet med CTD-kinase + helikase for smelting

37
Klasse II-gener transkribert av RNA polymerase II
TATA TFIID TBP TF RNAPII Mediator GTFs Chromatin Signal

38
PIC-dannelse stimuleres av transkripsjonsfaktorer
Dannelse av PIC styres av TFs som binder elementer oppstrøms Aktivatorer med høy sekvens-spesifisitet Rekrutteringspunkt for assembly av komplekser

39
(Oppstrøms) Transkripsjonsfaktorer
TAD DBD N C Modulær oppbygning - iallfall 2 domener: 1. DBD - DNA-bindingsdomene 2. TAD - transaktivatordomene DBD: grunnlaget for gruppering i TF-familier Sinkfingre, kjernereseptorer, helix-loop-helix, leucin-zippere,osv. TAD ulike grupper Tre klasser med ulike targets sure domener (GAL4, steroid receptor) glutamin-rike domener (Sp1) prolin-rike domener (CTF/NF1) Strukturløse i fri tilstand - 3D i kontakt med target?
 glutamin-rike domener (Sp1) prolin-rike domener (CTF/NF1) Strukturløse i fri tilstand - 3D i kontakt med target")
40
Mange ulike TFs Konstitutive TFs Regulatoriske TFs
Konstitutivt uttrykte faktorer som finnes i alle celler Binder cis-elementer lokalisert nær TATA/INR Eksempler CCAAT boks - binder ulike TFs (CTF/NF-I, CBFINF-Y) GC-rike bokser - binder Sp1 Regulatoriske TFs 1. Signal-responsive faktorer eks.: CRE, HSE, GRE - medierer respons overfor cAMP, heat shock, glucocorticoider 2. Celletypespesifikke faktorer
 GC-rike bokser - binder Sp1. Regulatoriske TFs. 1. Signal-responsive faktorer. eks.: CRE, HSE, GRE - medierer respons overfor cAMP, heat shock, glucocorticoider. 2. Celletypespesifikke faktorer.")
41
Konstitutive Sp1 binder GC-bokser
Noen eksempler

42
Synergi - mange faktorer stimulerer bedre enn én alene
Mange kontakter gir mer effektiv PIC assembly

43
Familier - oppdelt etter DBD struktur
Zinc finger bHelix-Loop-Helix (Max) STAT dimer Leucine zipper (Gcn4p) p53 DBD NFkB
 STAT. dimer. Leucine zipper. (Gcn4p) p53 DBD. NFkB.")
44
Familie Sinkfingerproteiner
TFIIIA fra Xenopus var første isolerte og klonede eukaryote TF Funksjon: aktivering av 5S RNA transkripsjon Primærstruktur TFIIIA Bygget opp som repetisjon 9x 30 aa minidomener + 70 aa unik region C-term Hvert minidomene konservert mønster av 2Cys+2His Hvert minidomene strukturert rundt et koordinert sink ion

45
Sinkfingerproteiner Sink koordineres av 2 Cys + 2 His
Antall fingre varierer: 2-37 Antall medlemmer uvanlig mange S.cerevisiae genomet: 147 C.elegans genomet 535 Flere hundre humane Zif gener

46
3D struktur av sinkfingre
Zif268 - første multi-finger struktur Hver finger et minidomene med -struktur major groove kontakt via -heliks i gjenkjenning av basetripletter

47
Kjerne-reseptorer - en sinkfinger-variant
To ”sinkfingre” utgjør en samlet enhet En finger kontakter DNA Dimeriserer ved binding til DNA

48
Kjernereseptorer Stor familie hvor DBD binder 2 Zn++ via tetraedrisk mønster av Cys Konservert DBD aa Medierer transkripsjonell respons på komplekse ekstracellulære signal klassiske steroid hormoner sekretert fra endokrine celler via blod målcelle diffusjon inn binder reseptor aktiveres modulerer målgener Evolusjonsmessig koblet til multicellulære organismer

49
Gal4p - enda en sinkfinger variant (6 Cys + 2 Zn)
")
50
Leucin-zipper - nesten glidelås
60-80 aa motiv funnet i en rekke dimere TFs Prototypiske eks.: GCN4, Fos, Jun, C/EBP, ATF, CREB mange mulige dimer-partnere gir tallrike kombinasjoner Dimer-dannelse via parallell coiled coil av a-helikser (ZIP) Hver 7.aa = Leu 3.5 aa pr turn (coiled coil) hver 7.aa lik posisjon Alle Leu samme side og dimerisering via “leucine zipper”
 Hver 7.aa = Leu. 3.5 aa pr turn (coiled coil) hver 7.aa lik posisjon. Alle Leu samme side og dimerisering via leucine zipper")
51
Leucin-zipper - nesten glidelås
Dimer via parallell coiled coil av a-helikser Hver 7.aa = Leu 3.5 aa pr turn (coiled coil) hver 7.aa lik posisjon Alle Leu samme side og dimerisering via “leucine zipper”
 hver 7.aa lik posisjon. Alle Leu samme side og dimerisering via leucine zipper")
52
Kontakter DNA som en pinsett

53
HLH - helix-loop-helix - lignende variant med brutte helikser

54
Klasse II-gener transkribert av RNA polymerase II
TATA TFIID TBP TF RNAPII Mediator GTFs Chromatin Signal

55
Koaktivatorer - brobyggere som også gir kromatin remodelering
remodellering

56
En rekke koaktivatorer og korepressorer deltar

57
Klasse II-gener transkribert av RNA polymerase II
TATA TFIID TBP TF RNAPII Mediator GTFs Chromatin Signal

58
Hvordan påvirker kromatin transkripsjon? .. som repressor
Chromatin maintains a restrictive ground state by blocking association between Pol II basic transcription machinery and DNA while permitting many activators to bind to their target sites. TFIID TBP Activator

59
Transkripsjonsfaktorer fremmer PIC dannelse + åpning av kromatin
Enzymes that covalently modify the nucleosomal histones ATP-dependent chromosomal- modifying enzyme

60
To måter å åpne kromatin på
1. Åpning av kromatin via styrt acetylering Histon-modifisering via HAT-aktivitet 2. Åpning av kromatin via styrt nukleosommobilisering ATP-avhengig prosess

61
Halene på histonene modifiseres som signaler til transkripsjon
Acetylation Methylation Phosphorylation Ubiquitination ADP-ribosylation

62
Topologi Selv åpnet kromatin er en topologisk utfordring for RNAPII

63
Hvordan kan modifisering av histoner påvirke kromatin?
Tradisjonell forklaring: Modifisering vil påvirke histonenes ladning, noe som igjen vil kunne gi en endret nukleosomstruktur og endrede egenskaper Nyere forklaring: Modifiserte aminosyrerester i histonene utgjør bindingsseter for andre proteiner som så avgjør kromatinets videre skjebne

64
Histonhalemodifikasjoner i eukromatin og heterokromatin
Models for euchromatic or heterochromatic histone tail modifications. Schematic representation of uchromatin and heterochromatin as accessible or condensed nucleosome fibers containing acetylated (Ac), phosphorylated (P), and methylated (Me) histone NH 2 -termini.
, phosphorylated (P), and methylated (Me) histone NH 2 -termini.")
65
Kombinasjoner av modifikasjoner forbundet med aktivt og inaktivt kromatin
Examples of combinatorial modifications in histone NH 2 -terminithat are likely to represent “imprints” for active or inactive chromatin.

66
Synergistiske og antagonistiske modifikasjoner i halen av H3 og H4
Proposed synergistic (connected arrowheads) or antagonistic (blocked oval line) modifications in histone H3 and H4 NH 2 -termini. The arrow with the scissors indicates possible proteolytic cleavage of the H3 NH 2 -terminus.
 or antagonistic (blocked oval line) modifications in histone H3 and H4 NH 2 -termini. The arrow with the scissors indicates possible proteolytic cleavage of the H3 NH 2 -terminus.")
67
Hvordan histonkoden oversettes
Fig. 2. Translating the “histone code”. Described protein modules of histone-modifying enzymes that have been shown to interact with site-specific methylation (chromodomain) or acetylation (bromodomain) marks in histone NH 2 -termini. A protein module that would selectively recognize phosphorylated positions is currently not known. Abbreviations: HMT, histone methyltransferase; HAT, histone acetyltransferase; HDM, histone demethylase; PPTase, protein phosphatase; HDAC, histone deacetylase.
 or acetylation (bromodomain) marks in histone NH 2 -termini. A protein module that would selectively recognize phosphorylated positions is currently not known. Abbreviations: HMT, histone methyltransferase; HAT, histone acetyltransferase; HDM, histone demethylase; PPTase, protein phosphatase; HDAC, histone deacetylase.")
68
Proteolytisk modell for fjerning av stabil metylering fra histon H3
Fig. 3. A proteolytic model to remove “stable” methylation marks from histone H3. Abbreviations: Ub, ubiquitin-conjugating activity; Ub protease, ubiquitin-directed proteolytic activity. Depending on the chromatin environment and/or the nature of the ubiquitin signal, a methylated H3 NH 2 -terminus may be removed by proteolytic processing (left; see also Fig. 1D), or the entire H3 molecule may be degraded (right).
, or the entire H3. molecule may be degraded (right).")
69
K9-metylering av histon H3 og HP1-protein i heterokromatin
Nucleosomes (blue spheres) are cross-linked by the heterochromatin-associated HP1 proteins (golden chains). The affinity of HP1 for heterochromatin is generated by Suv39h-dependent H3-K9 methylation (metal hook). Artwork by Hannes Tkadletz (IMP).
 are cross-linked by the heterochromatin-associated HP1 proteins (golden chains). The affinity of HP1 for heterochromatin is generated by Suv39h-dependent H3-K9 methylation (metal hook). Artwork by Hannes Tkadletz (IMP).")
70
Telomerer, heterokromatin og eukromatin
Packaging of yeast telomeres. Binding of RAP1 protein may be a key nucleating event in the assembly of these complexes. Complexes of SIR3 and SIR4 proteins bind to the hypoacetylated histones, extending along the DNA; SIR2 is also critical for stable maintenance of the silenced state. SIR2: NAD-dependent histone deacetylase

71
MeCP2 – et protein som bindes til metylert DNA
Bindes spesifikt til metylert DNA via et metyl-CpGG-bindende domene Rekrutterer transkripsjonsrepresjonskom-plekset mSin3A/HDAC Kan ”invadere” kromatin på en metyleringsavhengig måte og forskyve histon H1 fra kromatin in vitro Mutasjoner i mSin3A kan føre til Retts syndrom, en nevrologisk sykdom som stort sett rammer jenter Symptoms: Girls with Rett Syndrome appear to develop normally until 6 to 18 months of age. They then enter a period of regression, losing speech and hand skills they had acquired. Most girls develop seizures, repetitive hand movements, irregular breathing and motor-control problems. A slowing of the rate of head growth may also become apparent. The girls can live to adulthood, but most never regain the ability to use their hands or to speak.

72
Mulige måter å aktivere inaktivt kromatin på
Hypotetisk aktivering av inaktivt kromatin ved to mekanismer

73
Hypotetisk samarbeid mellom metylering og histonaktivering for opprettholdelse av aktivt/inaktivt kromatin

Liknende presentasjoner