Laste ned presentasjonen
Presentasjon lastes. Vennligst vent

1
KJB400 forelesning Voet & Voet Kapittel 29 + 33
RNA processering KJB400 forelesning Voet & Voet Kapittel

2
Kobling trx-trl Koblet translasjon /transkripsjon i prokaryoter
Adskilt i eukaryoter

3
Tre typer endringer Spalting, fjerning av sekvenser
Exo- eller endo-nukleolytisk spalting Splicing Påsetting av nukleotid(er) 5´-ende og 3´-ende Modifikasjon av spesifikke nukleotider
 5´-ende og 3´-ende. Modifikasjon av spesifikke nukleotider.")
4
Tre grupper RNA å modifisere
Pre-mRNA Påsetting Splicing Ribosomal RNA Spalting tRNA Modifikasjon

5
Pre-mRNA prosessering
Prokaryoter: primær transkript = mRNA Eukaryoter: transkripsjon/ translasjon adskilt, mRNA modifisert i kjernen før translasjon i cytosol

6
Transkripsjon - prosessering
cap AAAAAAAAAAAAA Pre-mRNA (hnRNA) mRNA Koblede prosesser
 mRNA. Koblede prosesser.")
7
Påsetting i 5´-ende: capping
Cap: 3 modifikasjoner 7-met-guanosin koblet til 5´-ende Kobling via 5´-5´trifosfat bro Skjer kotranskripsjonelt O2´-metylering av ribose Cap2, Cap1 (multicellulær), Cap0 (unicellulær) N6-metylering av adenine Cap-1 Cap-2
, Cap0 (unicellulær) N6-metylering av adenine. Cap-1. Cap-2.")
8
Enzymer involvert Capping skjer når RNA bare er baser lang - altså kotranskripsjonelt cap bindes til et ”Cap binding complex” CBC CBC stimulerer splicing og 3´-end prosessering 3 enzymer involvert 1.Trifosfatase fjerner et fosfat 2. Guanylyl transferase kobler på GMP 3. 7-metyltransferase modifiserer terminal guanosin Fosforylert CTD rekrutterer capping enzym

9
Modifisering av 3´- ende: poly-adenylering
Definert 3´-ende dannes ikke via terminering, men via prosessering Pre-mRNA heterogene 3´-ende, mRNA veldefinert 3´-ende Poly(A) haler påsettes i 3´-ende 20-50x A-strekk i en egen prosess dvs poly(A) ikke gen-kodet AAAAAAAAAAAAA cap
 haler påsettes i 3´-ende x A-strekk i en egen prosess. dvs poly(A) ikke gen-kodet. AAAAAAAAAAAAA. cap.")
10
Trimming av 3´-ende cap Upresis terminering Presis ende via
Presis ende via Spalting og polyadenylering AAAAAAAAAAAAA cap

11
Poly-adenylering - to-trinns prosess
Spalting nedenfor AAUAAA Innen 50 nt før en mindre konservert (G)U-rik Poly(A) hale lages av poly(A) polymerase Koblet: AAUAAA binder CPSF Cleavage and polyadenylation specificity factor Bundet CPSF stimuleres poly(A) polymerase
U-rik. Poly(A) hale lages av poly(A) polymerase. Koblet: AAUAAA binder CPSF. Cleavage and polyadenylation specificity factor. Bundet CPSF stimuleres poly(A) polymerase.")
12
Kotranskripsjonell prosessering

13
Prosessering i 3´-ende: Kotranskripsjonelle prosesser
Når RNAPII nærmer seg 3´-enden av transkriptet, skjer flere koblede prosesser Splicing av terminalt intron spalting ved poly(A)-site, påkobling av poly(A) hale, terminering nedstrøms for poly(A)-site og frigjøring av RNAPII Disse prosesser avhenger av CTD ”Cleavage-polyadenylation specificity factor” CPSF og ”cleavage stimulation factor” CstF binder spesifikt til CTD og finnes assosiert med holoRNAPII.
-site, påkobling av poly(A) hale, terminering nedstrøms for poly(A)-site og frigjøring av RNAPII. Disse prosesser avhenger av CTD. Cleavage-polyadenylation specificity factor CPSF og cleavage stimulation factor CstF binder spesifikt til CTD og finnes assosiert med holoRNAPII.")
14
Hvorfor poly(A)? Klippekort-hypotesen
PABP AAAAAAAAAAAAA Poly(A) beskytter mRNA mot degradering i cytosol Poly(A) bindes til PABP Poly(A) forkortes ettersom mRNA translateres cap AAAAAAAAAA cap AAAAAAA cap AAAA cap A cap ustabil
 beskytter mRNA mot degradering i cytosol. Poly(A) bindes til PABP. Poly(A) forkortes ettersom mRNA translateres. cap. AAAAAAAAAA. cap. AAAAAAA. cap. AAAA. cap. A. cap. ustabil.")
15
Splicing Kodende sekvens er i eukaryoter oftest stykket opp
avbrutt av ikke-kodende regioner Heterogen nukleær RNA hnRNA kb Større en protein skulle tilsi Rask turnover 1977: pre-mRNA har introns Som blir fjernet ved splicing

16
Eksempel: ovalbumin Presisjon - leseramme beholdes
Rekkefølge av exons beholdes Introns er som oftest større enn exons

17
Et typisk humant gen

18
Sekvens-signaler som definerer introns
Invariant GU i 5´-splice site (5´ss) Invariant AG i 3´- splice site (3´ss) Branch point sequence (BPS) Polypyrimidine tract (Py tract) Poly-Y tract Likevel så degenert at dataprogram bare klarer 50% treff i prediksjon
 Invariant AG i 3´- splice site (3´ss) Branch point sequence (BPS) Polypyrimidine tract (Py tract) Poly-Y tract. Likevel så degenert at dataprogram bare klarer 50% treff i prediksjon.")
19
Mekanisme:via 2 transesteri-fiseringer
Trinn 1 - dannelse av lasso-struktur (lariat) Exon-intron (5´) brudd og exon release Bro 2´-5´-fosfodiester A i forgrening: CURAY konsensus 20-50 foran 3´-splice site
 Exon-intron (5´) brudd og exon release. Bro 2´-5´-fosfodiester. A i forgrening: CURAY konsensus foran 3´-splice site.")
20
2´-5´-fosfodiester forgrening

21
Mekanisme:via 2 transesteri-fiseringer
Trinn 2 - fusjon av exoner Fri 3´-OH fra exon N danner fosfodiester binding med 5´-fosfat i exon (N+1) Avspaltet lariat-intron blir raskt degradert Uten fri energi input
 Avspaltet lariat-intron blir raskt degradert. Uten fri energi input.")
22
”Snurps” Hvert signal binder en snRNP
Small nuclear RNAs snRNAs binder protein og danner: Small nuclear ribonucleoproteins Disse gjenkjenner ulike splice-signaler, noen via base-paring

23
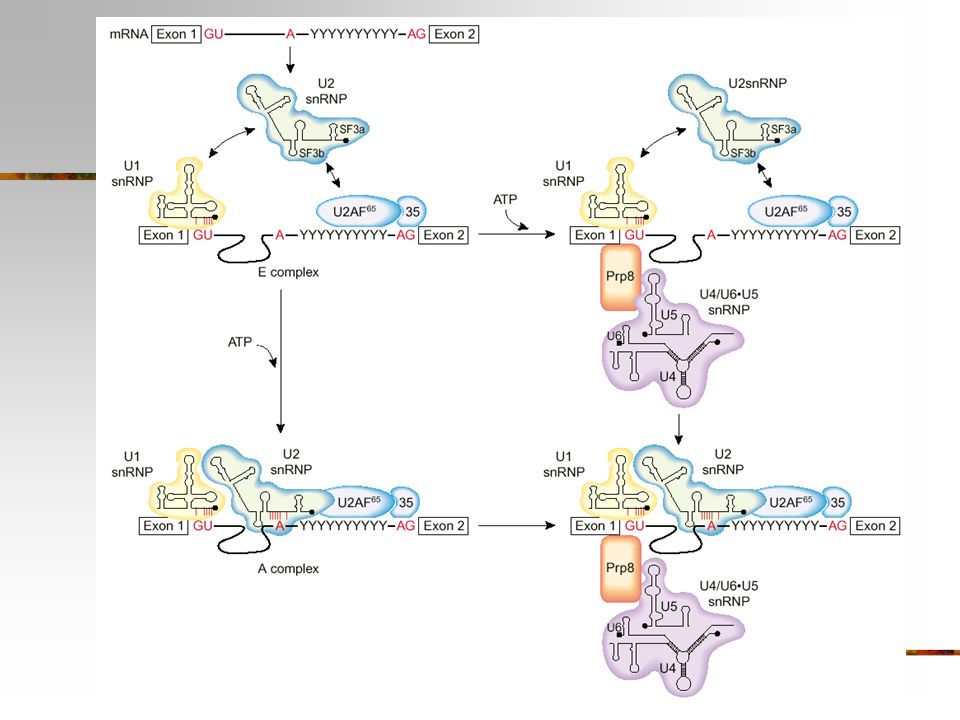
Spliceosomet utfører splicing
Spliceosomet = 5 snRNPs + prot = 50-60S U1, U2, U4, U5 og U6 snRNPs Mange andre non-snRNP proteiner Trinnvis assembly 5´ss bindes av U1 snRNP (E kompleks) Poly-Y + 3´ss bindes av U2AF Forgrenings-A bindes av U2 snRNP ATP-avhengig trinn U4/U6-U5 tri-snRNP assosieres og et kompetent splice kompleks dannes
 Poly-Y + 3´ss bindes av U2AF. Forgrenings-A bindes av U2 snRNP. ATP-avhengig trinn. U4/U6-U5 tri-snRNP assosieres og et kompetent splice kompleks dannes.")
25
Bro-dannelse: over exon og over intron
SR-proteiner involvert

26
Fortsatt mye ukjent

27
Hvorfor splicing? Genetiske fossiler eller … ….nyttig mekanisme?

28
Nytte: Alternativ splicing
En måte å øke protein diversitet uten å øke antall gener Drosophila Dscam genet genererer isoformer 576 alternativt splicede former av K+-kanal i fugleøre-reseptorer (rolle i gjenkjenning av lyd-frekvenser) Genom sml Humane genom bare gener Mer alternative splicing enn i lavere organism 3.2 alternative splice -former pr humant gen 1.34 alternative former pr gen i C.elegans
 Genom sml. Humane genom bare gener. Mer alternative splicing enn i lavere organism. 3.2 alternative splice -former pr humant gen alternative former pr gen i C.elegans.")
29
Eksempel: a-tropomyosin
7 Celletype-spesifikke varianter

30
Mange måter å variere på
Alternative 5´-splice sites (a) Alternative 3´-splice sites (b) Exon skipping/inclusion © Alternativ exon bruk (d) Intron retensjon (e)
 Alternative 3´-splice sites (b) Exon skipping/inclusion © Alternativ exon bruk (d) Intron retensjon (e)")
31
Alternative initieringsseter (alternative 1. Exons)
Alternative 1. Exons ≈ altern promotere Hvor separat regulering/nivå er nødvendig -amylase

32
Sykdommer pga splicing
Mange genetiske sykdommer skyldes feil splicing 15% av genetiske sykdommer er forårsaket av mutasjoner som ødelegger funksjonelle splicingsseter eller genererer falske nye

33
Exons ≈ proteindomener ?
W.Gilbert: exons svarer til primitive protein-domener som større proteiner er satt sammen av Eks. Pyruvat kinase

34
Evolusjonshypoteser: ”Intron early” eller ”Intron late”
Var der tidlig og er forsvunnet i prokaryoter Intron late Intron er blitt satt inn i intron-frie ORFs

35
Prosessering av ribosomal RNA

36
E.coli pre-rRNA prosessering
Endo- og exo-nukleaser

37
RNaser Endonukleaser: RNase III, RNase P, RNase E og F
Spalting i baseparede stems Exonukleaser: M16, M23 og M5 trimmer ferdig

38
rRNA er metylert N6,N6-dimetyl-adenin O2´-metylribose Funksjon ukjent
Beskyttende mot nukleaser som benytter 2´-OH

39
Eukaryot rRNA prosessering ligner den hos prokaryoter
Small nucleolar RNAs (snoRNA) involvert
 involvert.")
40
Prosessering og assembly av ribosomer

41
Prosessering av tRNA

42
tRNA - kløverblad-struktur
Mange baser er modifisert Antikodon 3´-CCA Kodet hos prokaryoter Appended hos eukaryoter via en tRNA nukleotidyltransferase

43
Splicing av tRNA introns
Noen eukaryote tRNAs har mini-intron Ved siden av antikodon

44
tRNA trimmes i 5´-ende av et ribozyme: RNase P
Rnase P = 377 nt RNA + 14 kD protein RNA katalytisk Protein (basisk) reduserer elektrostatisk frastøtning mellom ribozym og substrat
 reduserer elektrostatisk frastøtning mellom ribozym og substrat.")
Liknende presentasjoner




>")














